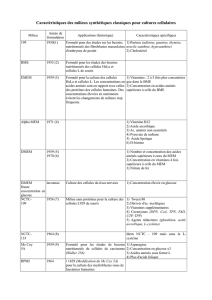
IUT A Génie biologique 2004 1 Année. JPPERRET

1
I.U.T. A Génie biologique
2004
1ère Année.
REGULATION HORMONALE et NERVEUSE du METABOLISME
ENERGETIQUE
J.P.PERRET

2
REGULATION HORMONALE et NERVEUSE du METABOLISME
ENERGETIQUE
( parties non développées en cours )
( parties non étudiées avec le groupe E )
1. Les substrats énergétiques : glucose, acides gras et acides aminés. p. 4
1.1: formes circulantes et de réserve et conditions d’utilisation :
glucose et glycogène ( musculaire et hépatique )
acides gras libres et triglycérides ( des adipocytes )
acides aminés et protéines ( musculaires )
1.2 :les tissus utilisateurs :
hématies ; neurones ; muscles et adipocytes ( insulino dépendants )
2 . Le glucose est nécessaire pour l’utilisation des substrats énergétiques : p. 6
3. L’ATP est formé extemporanément à partir de quatre processus principaux :
glycolyse anaérobie cytoplasmique indépendante de l’oxygène
ß oxydation,cycle de Krebs et phosphorylations oxydatives mitochondriales
dépendantes de l’oxygène
4. Le métabolisme glucidique chez les ruminants et les monogastriques : p. 8
4.1 : l’apport alimentaire du glucose
4.2 : le contrôle neuro-hormonal de la glycogénogenèse et de la glycogènolyse
musculaire et hépatique à l’état nourri et au cours du jeûne
4.3 : la néoglucogenèse et son contrôle neuro-hormonal
4.4 : au cours de la lactation
5. Le métabolisme lipidique chez les ruminants et les monogastriques : p. 13
5.1 : l’apport alimentaire d’acides gras
5.2 : le contrôle neuro-hormonal de la lipogenèse et de la lipolyse à l’état nourri
et au cours du jeûne
5.3 : la cétogenèse
5.4 : au cours de la lactation
6. Le métabolisme énergétique protéique p. 16
6.1 : l’apport alimentaire des acides aminés
6.2 : leur utilisation pour le métabolisme énergétique
7. Le contrôle de la sécrétion des hormones régulatrices du métabolisme énergétique p. 17
au cours du jeûne et à l’état nourri
7.1 : hormones pancréatiques : insuline et glucagon
7.2 : autres hormones : glucocorticoîdes
Exercices : p. 19

3
REGULATION HORMONALE et NERVEUSE du METABOLISME
ENERGETIQUE
Toutes les cellules d’un organisme utilisent en permanence des substrats énergétiques ( glucose
et acides gras surtout, mais aussi acides aminés ) pour assurer leur fonctionnement. L’utilisation de
ces substrats permet à chaque cellule de fabriquer extemporanément la seule source d’énergie qui
peut être transférée directement aux voies métaboliques : les 2 liaisons phosphate riches en énergie
de l’ATP.
La disponibilité des substrats énergétiques varie :
a) en fonction de l’ingestion des aliments,
b) de la qualité des aliments ingérés associée aux transformations qu’ils subissent dans le tube
digestif ainsi que
c) des capacités spécifiques de chaque type cellulaire à utiliser un substrat énergétique.
Aussi il est nécessaire qu’existent des mécanismes qui permettent de favoriser ou de limiter
chaque voie métabolique et plus précisément l’activité des enzymes qui y sont impliqués. Ces
mécanismes sont de trois types :
a) régulation de la synthèse et de la dégradation des enzymes par le contrôle de l’expression des
gènes, qui est surtout hormonal mais aussi nutritionnel.
a) régulation neuro hormonale de l’activité des enzymes par leur phosphorylation
b) régulation allostérique de l’activité des enzymes par un ou des substrats ou produits de la voie
métabolique dans laquelle ils sont impliqués.
Substrat
Régulation allostérique
( - )
régulation neuro hormonale (synthèse, activité) ENZYME
et nutritionnelle ( - )
Régulation allostérique
substrat transformé

4
Nous étudierons ici uniquement les régulations neuro hormonales des principales voies du
métabolisme énergétique. Cette étude nous permettra de différencier leur importance relative chez
les mammifères ruminants et non ruminants
1 Les substrats énergétiques : glucose, acides gras et acides aminés.
1.1: formes circulante et de réserve et conditions d’utilisation :
stockage limité = phosphagène ATP : production extemporanée = f (besoins)
ac. aminés glucose : (stockables) :ac. gras c. cétoniques ac. aminés (stockables)
- alimentation (après un repas)
- réserves corporelles (à jeun )
Origine des substrats énergétiques
- stockage du glucose = GLYCOGENE : foie et muscles
- stockage des acides gras = TRIGLYCERIDES : adipocytes
- ( stockage des acides aminés = PROTEINES : muscles )
Stockage des substrats énergétiques
La masse de glucose dans les liquides extracellulaires est d’environ 1g/l (0,5 chez les ruminants )
soit 0,15 g/kg de poids vif. Dans les cellules le glucose est immédiatement phosphorylé en glucose
6 phosphate ( G6P) avant d’être métabolisé ou stocké. Il est mis en réserve sous forme polymérisée,
le glycogène, dans deux tissus : le foie (environ1g/kg de poids vif, mais moins chez les ruminants )
et les muscles (environ 3g/kg de poids vif ).
Les destinées du glycogène hépatique et musculaires sont différentes. Le glycogène hépatique
est déstocké lors du jeûne ( c’est à dire lorsque le tube digestif est vide ) pour fournir du glucose qui
est sécrété dans le sang à l’usage des autres tissus. Cette réserve est épuisée après 18h de jeûne. Le
glycogène musculaire est déstocké lors du travail musculaire pour fournir du G6P utilisé
uniquement dans la cellule musculaire lorsque la quantité de glucose apportée par le sang au muscle
est insuffisante

5
La quantité d’acides gras circulant dans le sang sous forme libre ( ou non estérifiée = AGL ) est
d’environ 0,1 g/l. Dans les cellules les AGL sont utilisés après leur activation en acyl-CoA. Ils sont
métabolisés ou mis en réserve sous forme estérifiée de triglycérides dans le tissus adipeux blanc (
environ 120 g/kg de poids vif ). Les triglycérides représentent donc la réserve énergétique de loin la
plus importante, théoriquement épuisée après 60 jours de jeûne chez l’homme non obèse ( mais
seulement 6 chez le rat ) ce qui signifie que l’homme bénéficie d’un mécanisme de défense contre
le jeûne particulièrement efficace.
La quantité d’acides aminés circulant dans le sang est d’environ 0,2 g/l. Leur concentration
musculaire est 20 à 30 fois plus importante. Dans les cellules ils sont utilisés surtout pour la
synthèse protéique si l’alimentation est suffisante et équilibrée. En cas de jeûne prolongé ou
d’insuffisance alimentaire, certains d’entre eux sont utilisés comme précurseurs de substrats
énergétique : glucose ou corps cétonique. Les protéines musculaires représentent la réserve d’acides
aminés ( environ 90 g/ kg de poids vif ) utilisée dans ces cas.
1.2 : les tissus utilisateurs :
SUBSTRAT Cellules utilisatrices
Glucose uniquement
Glucose + corps cétoniques
Glucose + acides gras
Hématies
Neurones
Muscles et tissus adipeux(insulino dépendants)
et autres
Utilisation des substrats énergétiques
Les divers types cellulaires de l’organisme n’utilisent pas de la même manière les divers
substrats énergétiques.
Certaines cellules ne peuvent utiliser que le glucose. Ce sont les hématies.
D’autres n’utilisent habituellement que le glucose mais peuvent aussi utiliser, en cas de déficit de
glucose les corps cétoniques qui sont issus de la β oxydation des acides gras ou des acides aminés :
ce sont les neurones.
Les autres utilisent en proportion variable le glucose et les acides gras.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
1
/
21
100%