Effect of lactoferrin, probiotics and SM3 - Tel Archives ouvertes

Effect of lactoferrin, probiotics and SM3 (milk extract
enriched in sphingolipides) on immune function in mice
Cuibai Fan
To cite this version:
Cuibai Fan. Effect of lactoferrin, probiotics and SM3 (milk extract enriched in sphingolipides)
on immune function in mice. Life Sciences [q-bio]. AgroParisTech, 2008. English. <NNT :
2008AGPT0054>.<pastel-00005539>
HAL Id: pastel-00005539
https://pastel.archives-ouvertes.fr/pastel-00005539
Submitted on 3 Nov 2009
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-
entific research documents, whether they are pub-
lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destin´ee au d´epˆot et `a la diffusion de documents
scientifiques de niveau recherche, publi´es ou non,
´emanant des ´etablissements d’enseignement et de
recherche fran¸cais ou ´etrangers, des laboratoires
publics ou priv´es.

N° /__/__/__/__/__/__/__/__/__/__/
THÈSE
pour obtenir le grade de
Docteur
de
l’Institut des Sciences et Industries du Vivant et de l’Environnement
(Agro Paris Tech)
Spécialité : nutrition humaine
présentée et soutenue publiquement
par
FAN Cuibai
Le 22 septembre 2008
L’influence de la lactoferrine, de probiotiques et du SM3 (extrait enrichi en
sphingolipides) sur des fonctions immunitaires de la souris
Directeur de thèse : Daniel TOME
Travail réalisé : INRA-AgroParisTech, UMR914 Physiologie de la Nutrition et
du Comportement Alimentaire, F-75005 Paris
Devant le jury :
Mme. Christine HOEBLER, Ingénieur de recherche, INRA Nantes…………………...……Rapporteur
Mme. Isabelle OSWALD, Directeur de recherche, INRA Toulouse ………...………………Rapporteur
M. Saïd Bouhallab, Professeur, INRA Rennes.................................................................Examinateur
Mme. Anne-Marie DAVILA-GAY, Maître de Conférences, AgroParisTech ………….…Examinateur
Mme. Christine MARTIN-ROUAS, Responsable Nutrition, SOREDAB …...............…...Examinateur
M. Daniel TOME, Professeur, AgroParisTech …………………………………………..….Examinateur
L’Institut des Sciences et Industries du Vivant et de l’Environnement (Agro Paris Tech) est un Grand Etablissement dépendant du
Ministère de l’Agriculture et de la Pêche, composé de l’INA PG, de l’ENGREF et de l’ENSIA
(décret n° 2006-1592 du 13 décembre 2006)

Remerciements
Mes premiers remerciements iront à mon directeur de thèse, Daniel Tomé, qui m’a accueilli
au sein de son laboratoire.
Je tiens à exprimer ma profonde gratitude à Mr Michel Dubarry qui a su me faire bénéficier
de son expérience et de sa compétence. Je le remercie pour la confiance qu'il m'a toujours
témoignée en m'accordant une grande autonomie.
Un remerciement particulier à Mme Anne-Marie Davila-Gay pour ces corrections et ces
relectures averties. Elle a toujours su me consacrer son temps.
Je remercie aussi Mr François Blachier pour le temps qu’il a consacré à la correction de ma
thèse. Je le remercie aussi pour ces conseils et son soutien moral.
Je remercie tous particulièrement Mme Christine Hoebler et Mme Isabelle Oswald qui ont
accepté de juger ce travail et d'en être les rapporteurs.
Je remercie tous les membres de mon jury de thèse : Mme Christine Martin-Rouas, Mr Saïd
Bouhallab de me faire l’honneur d’assister à ma thèse.
Merci aussi à tous mes amis (e) et mes collègues du laboratoire qui se reconnaîtront ici. Je
leur exprime ma profonde sympathie et leur souhaite une bonne continuation.
Merci enfin pour ma famille pour leur soutien et leur patience au long de ces années de thèse.

SOMMAIRE
SOMMAIRE................................................................................................................ 1
RESUME.................................................................................................................... 5
ABSTRACT................................................................................................................ 6
ABREVIATIONS ........................................................................................................ 7
LISTE DE TABLEAUX............................................................................................... 8
LISTE DE FIGURES .................................................................................................. 9
INTRODUCTION GENERALE................................................................................. 10
SYNTHESE BIBLIOGRAPHIQUE ........................................................................... 12
I. Immunologie générale..................................................................................................................................... 12
I-1 Composantes de l’immunité ....................................................................................................................... 12
I-1.1 Immunité naturelle.............................................................................................................................. 12
I-1.2 Immunité acquise................................................................................................................................ 13
I-2 Organisation du système immunitaire ........................................................................................................ 13
I-2.1 Organes............................................................................................................................................... 13
I-2.1.1 rate............................................................................................................................................... 14
I-2.1.2 Les ganglions périphériques........................................................................................................ 15
I-2.2 Cellules du système lymphoïde........................................................................................................... 15
I-2.2.1 La lignée myéloïde...................................................................................................................... 15
I-2.2.2 La lignée lymphoïde.................................................................................................................... 18
I-2.3 Molécules impliquées dans système lymphoïde ................................................................................. 22
I-2.3.1 Immunoglobulines....................................................................................................................... 22
I-2.3.2 Le système du complément......................................................................................................... 24
I-2.3.3 Cytokines .................................................................................................................................... 24
I-3 Les sources de variation de la fonction immunitaire .................................................................................. 26
II. Ecosystème gastro-intestinal......................................................................................................................... 26
II-1 Le microbiote intestinal............................................................................................................................. 27
II-1.1 Distribution des bactéries dans le tube gastro-intestinal humain....................................................... 27
II-1.2 La flore de la souris ........................................................................................................................... 29
II-1.3 Etablissement de la flore.................................................................................................................... 29
II-1.4 Les principaux facteurs influençant le microbiote intestinal .............................................................30
II-1.5 Fonctions de la flore intestinale......................................................................................................... 30
II-2 La muqueuse intestinale ............................................................................................................................ 31
II-2.1 Les cellules épithéliales..................................................................................................................... 32
II-2.1.1 Les entérocytes .......................................................................................................................... 32
II-2.1.2 Les cellules caliciformes............................................................................................................ 33
II-2.1.3 Les cellules endocrines .............................................................................................................. 34
II-2.1.4 Les cellules de Paneth................................................................................................................ 34
II-2.2 Autres cellules de l’épithélium intestinal........................................................................................... 35
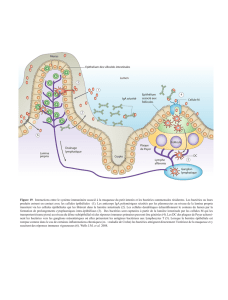
II-3 Système immunitaire associé à l’intestinal................................................................................................ 35
1

II-3.1 Cellules M et capture des antigènes................................................................................................... 36
II-3.2 Les plaques de Peyer ......................................................................................................................... 36
II-3.3 « homing » de cellules T.................................................................................................................... 36
II-3.4 Fonctions du système immunitaire intestinal (SII)............................................................................ 37
II-4 Relations des composants de l’écosystème gastro-intestinal..................................................................... 37
II-4.1 Le microbiote et l’épithélium intestinal............................................................................................. 38
II-4.2 Le microbiote et le système immunitaire........................................................................................... 39
III. Les aliments fonctionnels............................................................................................................................. 41
III-1 Lactoferrine.............................................................................................................................................. 42
III-1.1 Localisation et biosynthèse de la lactoferrine................................................................................... 42
III-1.2 Structure de lactoferrine ................................................................................................................... 43
III-1.3 Les récepteurs de la lactoferrine....................................................................................................... 44
III-1.4 Les fonctions biologiques de la lactoferrine..................................................................................... 45
III-1.4.1 Transport intestinal et devenir métabolique de la lactoferrine.................................................. 45
III-1.4.2 Transport du fer........................................................................................................................ 46
III-1.4.3 Effets antibactérien, antifongique et anti-viral de la lactoferrine.............................................. 47
III-1.4.4 Régulation de la myélopoïèse................................................................................................... 47
III-1.4.5 Modulation de la prolifération cellulaire .................................................................................. 48
III-1.4.6 Implication de la lactoferrine dans la réponse inflammatoire................................................... 48
III-1.4.7 Implication de la lactoferrine dans la réponse immunitaire...................................................... 49
III-2 Les probiotiques....................................................................................................................................... 51
III-2.1 Historique, développement et définition .......................................................................................... 51
III-2.2 Propriétés et critères de sélection des probiotiques.......................................................................... 52
III-2.3 Types de micro-organismes.............................................................................................................. 53
III-2.4 Connaissance en génétique............................................................................................................... 54
III-2.5 Persistance et survie des probiotiques dans l'environnement digestif .............................................. 54
III-2.6 Effets bénéfiques des probiotiques sur la santé................................................................................ 55
III-2.6.1 L’amélioration de la tolérance du lactose................................................................................. 55
III-2.6.2 Le traitement des infections gastro-intestinales........................................................................ 56
III-2.6.3 Les maladies inflammatoires de l’intestin ................................................................................ 56
III-2.6.4 La diminution du cholestérol sanguin....................................................................................... 57
III-2.6.5 La stimulation du système immunitaire.................................................................................... 57
III-3 Les sphingolipides ................................................................................................................................... 59
III-3.1 Structure des sphingolipides............................................................................................................. 60
III-3.2 Le métabolisme des sphingolipides.................................................................................................. 60
III-3.3 Les fonctions biologiques des sphingolipides .................................................................................. 61
III-3.3.1 Sphingolipides et signalisation cellulaire ................................................................................. 61
III-3.3.2 Sphingolipides et bactéries pathogènes .................................................................................... 61
III-3.3.3 Sphingolipides et cholestérol.................................................................................................... 62
III-3.3.4 Effets des sphingolipides sur la carcinogenèse......................................................................... 63
III-3.3.5 Sphingolipides et réactions inflammatoires.............................................................................. 64
MATERIEL ET METHODES .................................................................................... 65
I Animaux et aliments......................................................................................................................................... 65
I-1 Animaux ..................................................................................................................................................... 65
I-2 Condition d'élevage .................................................................................................................................... 65
I-3 Régimes ...................................................................................................................................................... 65
I-4 Suivi des animaux....................................................................................................................................... 66
II Analyses bactériologiques.............................................................................................................................. 66
II-1 Milieux pour le dénombrement des lactobacilles ...................................................................................... 66
II-2 Milieu pour le dénombrement des bifidobactéries .................................................................................... 67
II-3 Méthode de dénombrement des bactéries.................................................................................................. 67
II-4 Méthode de confirmation des genres Lactobacillus et Bifidobacterium ................................................... 68
II-4.1. PCR spécifique du genre Lactobacillus............................................................................................ 68
II-4.2. PCR spécifique du genre Bifidobacterium........................................................................................ 68
2
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
1
/
196
100%