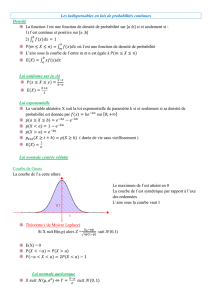
probabilité issu


Xnn
N
n
m Xn=m n
•n n + 1
•
n
m Qm= (qm
i, i ∈N)qm
i
m
i
•ηm= (ηm
j, j ∈N)
m ηm
jj
m
(Ω,A)
(X0,(Yn,m,k)n,m,k≥0,(Zn,m)n,m≥0).
X00Yn,m,k
k n
m Zn,m (n+ 1)
Xn=m
µN Pµ
(Ω,A)
•X0µ
•n, m, k Yn,m,k Qm
•n, m Zn,m ηm
X0Xn
Xn+1 =
m
X
k=1
Yn,m,k +Zn,m Xn=m, m ∈N,

0
Piµ i
X0=i
Xn+1 Xn(Yn,m,k, Zn,m, m, k ≥0)
(Xn)n
Pij =P(Xn+1 =j|Xn=i) = X
k1,k2,...,ki,r∈N,k1+···+ki+r=j
qi
k1qi
k2···qi
kiηi
r.
Qiηii
ηm
0= 1 ηm
0m
a= 1 a= 2
n X(1)
n1X(2)
n2
1 2 m1
1m22Qm1,m2
1= (Qm1,m2
1,i )i∈N
Qm1,m2
2= (Qm1,m2
2,i )i∈N
(Ya
n,m1,m2,k;n, m1, m2, k ∈N, a = 1 2)
Qm1,m2
aX(1)
0
1X(2)
02
a= 1 a= 2
X(a)
n+1 =
ma
X
k=1
Ya
n,m1,m2,k X(1)
n=m1, X(2)
n=m2, m1, m2∈N.

(X(1)
n, X(2)
n)N2
P(i1,i2)(j1,j2)=P(X(1)
n+1 =j1, X(2)
n+1 =j2|X(1)
n=i1, X(2)
n=i2)
=X
k1+···+ki1=j1;r1+···+ri2=j2
i1
Y
l=1
Qi1,i2
1,kl
i2
Y
m=1
Qi1,i2
2,rm.
1 2
2α1
α2(X(1)
n, X(2)
n)
P(i1,i2)(j1,j2)=X
A(i1,i2,j1,j2)
i1
Y
l=1
Qi1,i2
1,klµkl
k0
l¶αk0
l
1(1 −α1)kl−k0
l
i2
Y
m=1
Qi1,i2
1,rmµrm
r0
m¶αr0
m
2(1 −α2)rm−r0
m
A(i1, i2, j1, j2) ((kl, k0
l, rm, r0
m), l = 1,··· , i1;m=
1,··· , i2) 0 ≤k0
l≤kl0≤r0
m≤rmj1= (k1−k0
1) + ··· + (ki1−
k0
i1) + r0
1+··· +r0
i2j2=k0
1+··· +k0
i1+ (r1−r0
1) + ··· + (ri2−r0
i2)
(Xn)nn Xn→
0Xn
Xn→+∞(Xn)n
an0anXn
n

Qm=Q
ηm0Xn
n= 0,1,2, . . .
Xnn Yn,k
(1 ≤k≤Xn)
Xn+1 =
Xn
X
k=1
Yn,k (n≥0) .
Yn,k
YN
(qk, k ∈N)g(s) = E(sY) = Pkqksk
g(s)≡s
0 3 X0= 1 Y0,1=
2, Y1,1= 3, Y1,2= 1, Y2,1= 2, Y2,2= 0, Y2,3= 1, Y2,4= 3
X0= 1
X1= 2 X2= 4
X3= 6
.
 6
7
8
9
10
11
12
13
14
15
16
17
6
7
8
9
10
11
12
13
14
15
16
17
1
/
17
100%