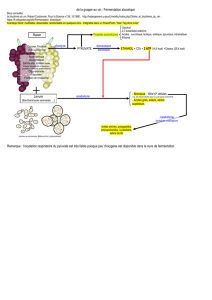
Communication présentée le 2 7 octobre 196

Communication présentée le 27 octobre 1962.
ETUDE DES RELATION S EX ISTANT ENTRE
LA SYNT H ESE DE S ACIDES AMINES
A PARTIR D O S E S ET DE DERIVES D ’O SES
ET LES PHENOM EN ES OSMOR EGULA TEU RS
AU NIVE AU DE NERFS ISOLES
DE H O M A R U S V U L G A R IS (L.)
p a r R. G i l l e s ,
S t a g ia ire d e re c h e r c h e s a u F .N .R .S .
I n s t i t u t L é o n F re d e ric q , B io c h im ie , U n iv e r s ité d e Liège.
Résum é. — L ’em ploi de glucose et de pyruvate marqués nous a permis
d ’aborder l’étude des mécanismes, peu connus, responsables de la varia
tion du pool intracellulaire des acides aminés libres en relation avec les
phénom ènes osmorégulateurs.
Les résultats obtenus semblent montrer qu’il existe, chez le homard,
une voie de form ation des acides aminés différente de la glycolyse, uti
lisant le glucose, voie m étabolique nécessitant de surcroît l’utilisation
de l ’alanine.
L’étude de l'influence des facteurs amenant une modification de l’équi
libre cationique intracellulaire (vératrine, cocaïne et stim ulation) semble
indiquer que la composition du pool intracellulaire des acides am inés
libres est sous la dépendance étroite de la composition cationique de la
cellule par une variation de l ’équilibre dégradation - synthèse des acides
aminés.
F l o r k i n et ses collaborateurs o nt dém ontré que la variation
du pool in tracellulaire des acides am inés libres pouvait être mise
en relation avec l'osm orégulation du liquide intracellulaire chez
les invertébrés euryhalins ( D u c h a t e a u e t F l o r k i n , 1955: D u c h a -
t e a u e t F l o r k i n , 1 9 5 6 ; J e u n i a u x , B r i c t e u x e t F l o r k i n , 1 9 6 1 ... ) .
N éanm oins, peu de choses sont connues qu an t au x m écanismes
responsables des variations observées. Il a été suggéré que la
composition cationique du liquide intracellulaire p ourrait jouer
un rôle im portant dans la com position de ce pool ( S c h o f f e n i e l s ,
i9 6 0 ).
N ous avons abordé l’étude de ce problèm e en exa m ina nt les
relations existant en tre le m étabolism e des oses et dérivés d ’oses
et la synthèse des acides aminés dans diverses conditions expé
rim entales am enant une m odification de l’équilibre cationique

R . G IL L E S
intracellulaire. L'action de la Stim ulation et de certains agents
pharm acodynam iques (la vératrine et la cocaïne, p ar exemple)
pou vant s’interpréter en term e de m odification de l’équilibre
cationique intracellulaire ( W o l l e n b e r g e r , 1955; K ini e t Q u a s t e l ,
i960), nous avons étudié l ’incorporation de 14C dans les acides
aminés à p artir de glucose-U-14C et de pyruvate-1-14C dans ces
diverses conditions expérim entales au niveau de la chaîne n er
veuse du hom ard.
Si nous comparons les résultats obtenus avec les témoins en
utilisan t le glucose et le pyruvate com m e substrat, nous rem ar
quons que les acides aminés don t la synthèse s’effectue à partir
d ’interm édiaires du cycle de Krebs sont m arqués égalem ent,
que le sub strat utilisé soit le glucose ou le pyruvate. Cette cons
tatatio n peut s’expliquer en postulant que le pyruvate en tra n t
dans le cycle de Krebs, se divise en deux lots quantitativem ent
de mêm e importance. L a première moitié d u pyruvate entre dans
le cycle de K rebs sous forme de m alate et, de ce fait, ne subit
pas de décarboxylation. Cette interprétation im plique donc
q u ’une m oitié seulement du p yruvate-i-14C entran t dans le cycle
de Krebs garde son activité spécifique. Or, cette activité est dou
ble de celle du pyruvate provenant de la dégradation du glucose
exogène puisque, au d épart des expériences, l’activité spécifique
des deux substrats utilisés est la m ême et que la dégradation
d'un e molécule de glucose p ar la voie du cycle d ’Em bden-
M eyerhof fournit deux molécules de pyruvate. Il résulte donc
de ceci que l’activité spécifique du pyruvate entran t dans le cycle
de K rebs sera la m ême quel que soit le substrat exogène utilisé
et dès lors, quel que soit ce substrat, l’activité des acides am inés
dicarboxyliques sera du m ême ordre de grandeur.
Ces résultats m on tren t de plus que, en ce qui concerne l’ala-
nine, l’activité est toujours plus élevée lorsque le substrat utilisé
est le pyruvate-1-1'C. Cependant, les différences d ’activité obser
vées ne sont pas de l ’ordre de celles attendues, en considérant
que l’activité spécifique du pyruvate provenant de la dégradation
du glucose exogène est deux fois m oindre que celle obtenue dans
les expériences utilisan t directem ent le pyruvate com m e substrat.
E n effet, les différences d ’activité observées ne sont pas de l ’or
dre de 2, m ais de l’ordre de 4. Il semble donc q u ’il existe, chez
le ho m ard tou t au moins, une voie métabolique différen te de la
glycolyse utilisant le glucose, voie m étabolique nécessitant de
surcroît l’utilisation de l’alanine.
L ’étude de l’influence des trois facteurs précédem m ent cités
(stim ulation, vératrine et cocaïne), d ont l’action est interprétée

O SM O REG U LA TIO N DANS L E S N E R F S DE H O M A R D 19 3
comm e é tan t une m odification de l’équilibre cationique intra
cellulaire, sur la synthèse des acides aminés à p a rtir doses et de
dérivés d ’oses, m ontre que cette synthèse est sujette à des varia
tions. E n effet, les résultats obtenus m ontrent que l’add ition de
sulfate de vératrine 4 ¡J.M au m ilieu incubateur amène l ’augm en
tation de la synthèse de tous les acides aminés étudiés à l’excep
tion de la lysine et de l’arginine. Il en est de m ême en ce qui
concerne la stim ulation électrique prolongée. P ar contre, les
résultats obtenus p ar action du chlorure de cocaïne 4 u.M font
état de la dim inution de la synthèse des acides am inés dicarbo-
xyliques.
En conclusion, il semble donc que la composition d u pool
intracellulaire des acides aminés libres soit sous la dépendance
étroite de la compositon cationique de la cellule, pa r une varia
tion de l’équilibre dégradation - synthèse des acides am inés.
Su m m a r y .
T h e use of labelled glucose and p yru vate has pe rm itted th e stu dy of
th e little known m echanism s responsible fo r th e variation of th e in tra
cellular pool of free amino-acids in relation to osmoregulation.
In th e lobster, th e results seem to show tha t th ere is a path way using
glu cose fo r the form ation of amino -acids which is differen t from th e
glycolysis. M oreover, this pathw ay w oidd n eed alanine.
T h e stu dy of th e influence of factors leading to a change in the in tra
cellular cathionic equilibrium (veratrine, cocaine a n d stim ula tion) seem s
to show th a t th e com position of the intracellular p ool of free amino-
acids d epends on the cathionic com position of the cell by a variation
of the degradation - synthesis equilibriu m of the am ino-acids.
BIBLIOGRAPHIE.
DUCHATEAU, G h . e t F L O R K IN , M. (1955). — C o n c e n tra t io n d u m ili e u
e x té r ie u r e t é ta t s ta ti o n n a i r e d u po o l des a c id e s a m in é s n o n p r o t é iq u e s
des m u sc le s á 'E rio c h eir s in e n sis (M. E dw .). A rch, in te r n a t. P h y s iol. B io c h .,
63, 249.
DUCHATEAU, G h . e t F LO R K IN , M. (1956). — S y stè m es in tr a c e l lu l a ire s
d 'a c id e s a m in é s e t o s m o ré g u la t io n d es C ru sta c é s. J . d e P h ysio l.. 48, 520.
JEUN IA U X , Ch., B R IC T E U X , S. e t FLO R K IN , M. (1961). — C o n tr i b u tio n
d e s a c id e s a m in é s lib re s à l a r é g u la tio n o s m o tiq u e i n tra c e l l u la ire c h e z
d e u x C r u s ta c é s e u r y h a lin s L ea n d e r ser r a tu s F. e t L e a n d e r s q u illa L. C a h .
d e B io l. m a r., 2, 373.
K I N I, M. M. a n d Q U ASTEL, J . H. (1960). — E f f e c ts o f v e r a tr in e a n d c o cai
n e o n ce re b ra l c a rb o h y d r a te -a m in o a c i d s in te r re la t io n s . S c ie n c e , 1 3 1 , 412.
SC H O FFE N IELS, E. (1960). — O r ig in e d e s a c id e s a m in é s in te r v e n a n t d a n s
la r é g u l a tio n d e la p re s s io n o sm o tiq u e i n tra c e llu l a ire d e E rio c h eir s i n e n
sis M. Edw. A rch, i n te r n a t. P h y siol. B io c h ., 68, 696.
W O L LEN B E R G E R , A. (1955), — A c tio n o f p ro t o v e r a trin e o n t h e m etab o lism
of ce r e b ra l co rte x . B io ch . J., 6 1 , 68.
Institut Léon Fredericq, Biochimie.
Université de Liège.
1
/
3
100%