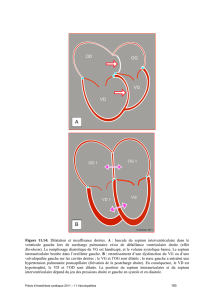
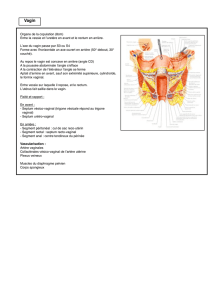
Embryologie cardio-vasculaire

1
Cardiologie
Mme Délézoïde
Le 13/02/08 à 15H30
Ronéotypeuse : Julie Seroussi
Embryologie
cardio-vasculaire

2
PLAN DU COURS
I. INTRODUCTION
II. FORMATION DES TUBES ENDOCARDIQUES PRIMITIFS ET
DETERMINATION DU SITUS VISCERAL
III. DELIMITATION : BASCULE CENTRALE DES TEP
IV. TUBES CARDIAQUES PRIMITIFS AU SEIN DE L’EMBRYON
V. HISTOGENESE
VI. EVOLUTION DU TUBE CARDIQUE
1. Allongement
2. Plicature
3. Convergence
VII. CLOISONNEMENT
1. Cloisonnement du sinus veineux
2. Cloisonnement du canal atrio-ventriculaire
3. Cloisonnement de l’oreillette primitive
4. Cloisonnement du ventricule primitif

3
I. INTRODUCTION
Le système cardio-vasculaire est un système qui va devoir se mettre en place très vite. En
effet, les réseaux vasculaires (aussi bien le réseau intra embryonnaires que les réseaux
extra embryonnaires) se mettent en place très tôt au cours du développement. Le cœur
devra également se mettre en fonction rapidement car il a une fonction de pompe et va
donc permettre de faire circuler le sang dans les vaisseaux et ainsi d’oxygéner les tissus.
Le développement cardiaque est donc un développement précoce et rapide. Il va
commencer vers J16 avec la gastrulation et à J50 pratiquement, c’est-à-dire avant la fin de
la période embryonnaire, le cœur sera complètement terminé : il aura 4 cavités et ce
jusqu’à la fin de la grossesse.
L’originalité du développement cardiaque est que pendant que le cœur va se développer le
sang va déjà circuler, exister dans les conduits vasculaires. De J21 à J 50 (c’est à dire
pendant l’essentiel du développement cardiaque) le cœur aura déjà sa fonction de pompe
et sera déjà le siège d’un flux sanguin. Et ce flux sanguin va jouer un rôle majeur dans le
développement du cœur. C’est l’un des exemples de l’intervention de la mécanique des
fluides au cours du développement d’un organe.
Beaucoup de malformations cardiaques sont liées à des perturbations du flux sanguin
intra cardiaque et ces perturbations vont elles-mêmes induire par exemple le défaut de
développement d’une cavité par rapport à une autre.
Ces cardiopathies in utero peuvent évoluer c’est-à-dire que d’une forme peu sévère au
départ elles peuvent devenir très sévères et conduire à la mort de l’enfant à la naissance ou
à une prise en charge chirurgicale immédiate.
Le cœur est donc un organe extrêmement dépendant de la mécanique des fluides
(mécanique du flux sanguin) et toute perturbation de ces flux perturbe le développement
cardiaque. Ce développement cardiaque épouse un peu ces flux sanguins et va se
poursuivre tout au long de la vie intra utérine.

4
J16 (gastrulation) à J50 (cœur «4 cavités »)
1
ère
étape du développement du cœur :
Lors de la gastrulation on a un ensemble de cellules ectoblastiques qui va migrer par la
ligne primitive et le nœud de Hansen pour venir se placer entre l’ectoblaste et l’entoblaste.
Ces cellules vont revêtir l’ensemble de la surface embryonnaire sauf au niveau des
membranes pharyngienne et cloacale. Elles vont constituer le mésoblaste.
Les cellules, qui au cours de la gastrulation viennent peupler l’interstice entre l’ectoblaste
et l’entoblaste au niveau de l’aire cardiaque, juste en avant de la membrane
pharyngienne, sont des cellules mésoblastiques qui ont un aspect mésenchymateux.
Ces cellules sont déjà extrêmement différenciées, c’est-à-dire que déjà alors qu’on a un
embryon qui est morphologiquement complètement symétrique, elles savent très bien
s’organiser. Elles savent où est la droite et la gauche de l’embryon, où est la partie
céphalique et caudale. Elles sont déjà déterminées pour fabriquer du cœur, c’est à dire
pour fabriquer d’une part un endocarde et d’autre part un myocarde.
Elles expriment déjà des gènes, facteurs de transcriptions qui vont permettre la
différenciation dans ce sens là :
NKX2-5 et GATA4 : codent pour des facteurs de transcription qui sont majeurs dans le
développement cardiaque.
Ces cellules sont donc déjà organisées dans tous les sens de l’espace.

5
Ces premières cellules qui arrivent à la gastrulation sont des cellules mésoblastiques
standards qui constituent le premier champ cardiaque. Une deuxième et troisième source
de cellules participeront à la formation du cœur.
A 50 jours l’embryon a un cœur à 4 cavités qui fonctionne parfaitement.
II. FORMATION DES TUBES ENDOCARDIQUES PRIMITIFS ET
DETERMINATION DU SITUS VISCERAL
Lors de la gastrulation, les cellules qui passent par la ligne primitive sont déjà orientées à
cause de ce qui se passe au niveau du nœud de Hansen pour savoir quel est le côté droit et
quel est le côté gauche. Il y a des cellules ciliées qui battent au niveau du nœud de Hansen
et qui vont envoyer du FGF vers la gauche et donc on va avoir une expression de SHH du
côté gauche de l’embryon et on a l’expression de récepteurs à l’activine du côté droit du
nœud de Hansen. Cette expression à gauche de SHH va induire dans le mésenchyme
l’expression de tas de gènes qui sont des «gènes de la gauche » tandis qu’il y aura d’autres
gènes qui vont s’exprimer dans le mésenchyme à droite qui seront des «gènes de la
droite ».
Tous ces facteurs de transcription vont donc agir sur des gènes pour modifier la matrice
extra cellulaire, l’activité mitotique, le cytosquelette des cellules… Et c’est cela qui va
induire toute cette transformation de l’embryon.
La latéralité joue un rôle majeur dans le développement du cœur. Si ce déterminisme
droite / gauche se fait mal, le cœur sera un des premiers organes à s’exprimer sous forme
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
1
/
24
100%