X fragile quoi de neuf - iPubli

MINI-SYNTHÈSE
8:
fragile quoi de neuf
=

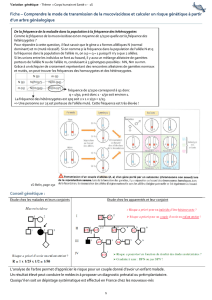
Nombre de répéons
de sque

?
FRM-1
FMR -1
FMR -1
notion de seuil
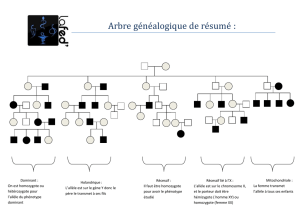
Hélène Gilgenntz
In
.
X N
Med
X Ce
et
X Ce
X
Ce
X
H
X
Med G
X
Med G
l.
1
/
3
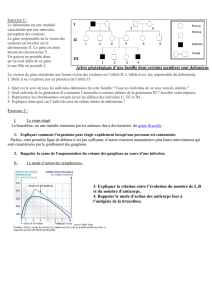
100%