photosynthese et metabolisme carbone

1
PHOTOSYNTHESE ET METABOLISME CARBONE
Le stomate représente la structure de régulation par détermination de la quantité de CO2 qu’il
importe et d’H2O qu’il exocyte.
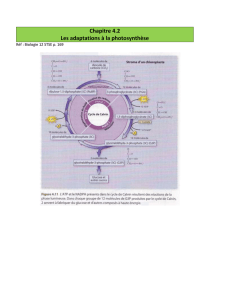
I/Le cycle photosynthétique de réduction du carbone (CPR).
1) Mise en évidence du cycle de réduction du CO2.
Montage expérimental de Melvin Calvin (prix Nobel de chimie en 1961) :
- Introduction de 14CO2.
- Arrêt brusque à t souhaité par ajout d’éthanol.
- L’éthanol sert comme solvant pour la chromatographie sur papier.
Sur la chromatographie on a présence de molécules intégrées par le 14CO2.
Premier produit formé : acide 3-phosphoglycérique (APG), 14C au niveau de la fonction acide.
Accepteur du CO2 : ribulose 1,5 biphosphate (RuBP).
2) Le cycle de Calvin.
Trois phases :
- Fixation du CO2 par carboxylation du RuBP.
- Réduction de l’APG.
- Régénération du RuBP.
La carboxylation est catalysée par la RubisCO :
RuBP + CO2 2 APG
La réduction de l’APG se fait par :
Hydrolyse de l’ATP.
APG + ATP ABPG + ADP
Oxydation du NADPH.
ABPG + NADPH GAP + NADP+
Assimilation du CO2.
3) Régulation du cycle de Calvin.
a) La RubisCO :
La RubisCO est une enzyme de régulation qui elle-même est régulée par la lumière et ses
intermédiaires :
- CO2 : activateur de la RubisCO fixation au carbamate.
- Mg2+ : Le Mg2+ sort du lumen par gradient de protons inverse pour activer le carbamate par
fixation de l’ion.
- RubisCO activase : elle active la RubisCO par libération du RuBP qui lui est fixée (nécessite
hydrolyse d’ATP).
Toutes ces réactions dépendent du pH et du gradient de protons de la photosynthèse.
Le CAIP est un inhibiteur de la RubisCO qui est dégradée par la lumière.

2
b) Autres enzymes de régulation :
La FBPase :
- Activation par la lumière bloquée par DCMU.
- Activation par la lumière bloquée par pont triol.
- Implication par ferrédoxine et thiorédoxine.
- Enzyme activée à l’obscurité par apport d’un agent réducteur.
c) Régulation par le phosphate inorganique :
S’il y a trop de DHOAP, il sera utilisé par l'enzyme pour faire du saccharose.
Si le Pi s'accumule, cette enzyme sera inhibée, permettant de dégrader de l'amidon en DHOAP.
II/La RubisCO.
La RubisCO :
- 50% des protéines du stroma.
- Héxadécamère de deux sous-unités.
L codée par le génome chloroplastique.
S codée par le génome nucléaire.
La RubisCO existe sous deux formes :
- Active : le jour car carbonylée.
- Inactive : la nuit car fixée au ribulose.
Le CO2 est un catalyseur, deux molécules sont nécessaires pour activer la RubisCO, la première fixe
le Mg2+ la seconde sert de substrat à la réaction.

3
III/Fonction oxygénase de la RubisCO.
Ce processus respiratoire n’a lieu qu’en présence de lumière et consomme de l’O2 en relâchant du
CO2 au sein du chloroplaste : photorespiration.
Ce processus a été démontré par utilisation d’isotopes radioactifs de l’O2.
L’activité oxygénase diminue de 20% l’activité de la RubisCO.
En consommant l’oxygène elle évite ainsi la création de dérivés oxygéniques nocifs pour la plante.
Le processus aboutit à la formation d’une molécule en C5, d’APG et d’acide glycolique (C2).
Le cycle en C2 fait intervenir :
- Chloroplaste.
- Peroxysome.
- Mitochondrie.
Fonctionnement du cycle :
I. Sortie de glycolate du chloroplaste.
II. Oxygénation du glycolate dans le peroxysome où la fonction alcool devient aldéhyde.
III. Formation de glycoxylate (génère H2O2 qui est dégradé par les catalases).
IV. Formation de glycine par transamination du glycoxylate qui est exportée vers la
mitochondrie.
V. Formation de Sérine par consommation d’un NAD+ à partir de la glycine.
VI. Formation d’hydroxypyruvate par perte du groupement amine.
VII. Formation de glycérate par consommation de NADH.
VIII. Renouvellement du RuBP par cycle CPR et consommation d’ATP.

4
La photorespiration ou cycle en C2 est corrélée au cycle de Calvin, le CO2 sortant du cycle en C2 est
intégré au cycle de Calvin. Selon la quantité de substrat dissout (O2 ou CO2) disponible, un des
deux cycles sera privilégié.
IV/La synthèse des produits terminaux.
Une grande concentration de triose phosphate bloque le processus, ils sont transportés hors du
stroma par antiport avec le Pi.
Les trioses phosphates (DHOAP) sont déphosphorylés et isomérisés en glucose.
Production des hexoses phosphate qui génèrent du saccharose.
Le transport des trioses phosphate est régulé par une transformation transitoire en amidon :
- Evite le blocage du transport.
- Augmente la pression osmotique (triose polymère).
Bilan du cycle :
Il faut 3 ATP et 2 NADPH par carbone incorporé, le bilan énergétique peut s'écrire :
CO2 + H2O CH2O + O2 ΔG= 480 kJ
2 NADPH, H+ + O2 2 NADP + 2 H2O ΔG= -440kJ
3ATP + 3 H2O 3 ADP + 3 Pi + 3 H+ ΔG= -92 kJ
Le bilan énergétique est excédentaire d'environ 50 kJ réaction favorisée.
V/Métabolisme des plantes en C4 et CAM.
Chez certaines plantes, la première molécule acceptant le CO2 est en C4 et non en C3.
1) Carboxylation supplémentaire.
Les mécanismes en C3 sont toujours présents malgré une seconde carboxylation qui se fait en
amont.
Fonctionnement du mécanisme :
I. Le système enzymatique PEPc agit sur le PEP (C3) par fixation d’un CO23
II. Formation d’une première molécule en C4 : oxaloacétate.
III. Deux voies se différencient pour la suite du processus :
Réduction de l’OAA en malate par une malate déshydrogénase
NADP+ dépendante.
Transamination de l’OAA en glutamate ou aspartate à l’aide d’une
aminotransférase.
Le PEP utilise le CO2 sous forme dissoute (HCO3
-) car son Km est beaucoup plus faible que celui de
la RubisCO.
Le but de ce processus est de libérer du CO2 au voisinage de la RubisCO ce qui diminue fortement
sa fonction oxygénase et réduit voir annule le manque apporté par la photorespiration.
Le fonctionnement successif de ces deux systèmes est dû à une anatomie particulière.
2) Morphologie générale.
Deux types cellulaires présents :
- Mésophile : lieu de la première carboxylation par le PEPc.
- Cellule de la graine périvasculaire (chloroplaste sans granas) : lieu de la seconde carboxylation.

5
L’OAA relâche du CO2 dans le cytoplasme de la cellule périvasculaire ce qui augmente le rapport
CO2/O2 et limite la photorespiration.
Le CO2 libéré est utilisé dans le cycle de Calvin.
Le transport entre les deux cellules :
- Se fait par voie symplastique (favorisée par une paroi suberifiée).
- L’OAA est réduit en malate par le NAD+.
- Le malate peut traverser le plasmodesme et est décarboxylé en pyruvate dans la gaine
périvasculaire.
- Transport du pyruvate à la cellule mésophillienne ou il est phosphorylé en PEP (consommation
en ATP).
Séparation spatiale des cycles.
La structure spéciale du chloroplaste de la graine périvasculaire possède peu de PSII diminuant la
lyse de l’eau et la libération d’O2, le rapport CO2/O2 augmente.
Les différentes variantes de cycle :
- Cycle de régénération du malate NAD+ dépendant.
- Cycle de régénération de l’aspartate NAD+ dépendant (minoritaire).
- Double navette du PEP.
3) Régulation de la voie en C4.
Le PEPc existe sous forme :
- Active : phosphorylé.
- Moins active : déphosphorylé.
La phosporylation est catalysée par une PEPc kinase en présence de lumière et la
déphosphorylation est catalysée par une phosphatase à l’obscurité.
Régénération du PEP :
Pyruvate + ATP + Pi PEP + AMP + PPi
Système par phosphorylation (obscurité) d’une enzyme PPDK (impliquée dans la régénération du PEP
par consommation d’ADP) qui catalyse AMP + ATP 2 ADP et PPi 2Pi.
En présence de lumière la PPDK est activée par déphosphorylation.
Régulation par le malate : PEPc non phosphorylé inhibé à l’obscurité par de faible
concentration du malate.
4) Différences plantes C3 et plantes C4.
Plantes
C3
C4
Saturation CO2
Lente
Rapide
Compensation CO2
Bas
Haut
Saturation lumière
Rapide
Lente
Compensation lumière
Bas
Haut
Rendement photosynthèse
Rapide et faible
Lent et fort
Efficacité température
25°C
35°C
 6
6
1
/
6
100%