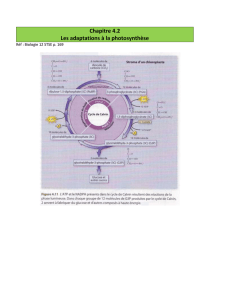
II) Etude du cycle de Calvin On peut résumer en 3 étapes le cycle de

II) Etude du cycle de Calvin
On peut résumer en 3 étapes le cycle de Calvin et Benson
1ere étape : l’incorporation du cO2 dans le ribulose-1,5-bisphosphate
2e étape réduction de l’APG (acide phosphoglycérite) en glycéraldéhyde-3-phosphate
3e étape : régénération
Premièrement le CO2 atmosphérique se fixe à un accepteur en C5 qui est carboxylé (RUBP ou
ribulose-1,5-bisphosphate). Il y a formation d’un intermédiaire en C6 instable, aussitôt hydrolysé en 2
molécules d’acide-3-phosphoglycérique (APG ou 3 phosphoglycérate). C’est le première
intermédiaire stable du cycle de Calvin. L’enzyme qui intervient est la plus importante du monde : la
rubisco
Deuxièmement réduction de l’APG en trioses phosphate. Dans le stroma du chloroplaste, l‘APG est
réduit à la lumière en aldéhyde phosphoglycérique (aldPG ou glyceraldéhyde-3-P). cette réduction se
réalise en 2 étapes, catalysées par la phosphoglycérokinase et pas la glycéraldéhyde-P-
désydrogehase. Au total, la formation de deux molécules de triose consécutives à la fixation d’une
molécule de CO2 nécessite deux molécules d’ATP et deux molécules de NADPH
Troisièmement la régénération du ribulose-1,5-bisphosphate (RUBP)
Le cycle de Calvin permet la régénération du RUBP à partir d’une partie des trioses formés. Ceci va
permettre l’incorporation de nouvelles molécules de CO2. Une partie de l’aldéhyde
phosphoglycérique va être utilisé pour régénérer l’accepteur en C5.
Approche rapide :
1 C5 + 1 CO2 (1 C6) 2 C3,
1 C3 synthèse du saccharose (cytoplasme) ou amidon (chlorolaste).
Pour régénérer le C5 soit à partir d’un C3, possible que si l’on utilise l’équilibre suivant : 5 C3 _ 3 C5,
Le cycle de Calvin se résume donc aux équations suivantes :
3 C5 + 3 CO2 (3 C6) 6 C3 5 C3 + 1 C3 3 C5 + 1 C3
III- Bilan du cycle photosynthétique
On a:
6CO2 +6 ribulose 1,5bisP 12 acide P glycérique
12 acides P glycérique + 12 ATP + 12 NADPH + 12 H+ 12 C3-P +12 ADP +12 NADP++12 Pi
10C3-P +6 ATP 6 rib-1,5-BisP +6 ADP+ 4Pi
2C3-P [hexose] + 2Pi
6CO2 + 18 ATP + 12NADPH + 12 H+ [hexose C6 H 12 O 6 ] + 18 ADP +18Pi +12NADP+
IV- Régulation du cycle de Calvin
La lumière va agir indirectement sur l’activité de certaines enzymes. L’activité des enzymes est
fortement liée au pH. A la lumière il y a alcanalisation du stroma. Le pH passe à 7,8. Il y a
augmentation des teneurs en mg2+. Il y a apparition de réducteur comme la feredoxine réduite.
Certaines ne sont fonctionnelles qu’à des pH légèrement basique. La lumière n’a donc pas pour seule

fonction de produire de l’ADP (ATP ???)et du NADPH, elle joue un rôle dans le fonctionnement du
chloroplaste. A l’obscurité on observe les phénomènes inverses.
V- Synthèse du saccharose
Il y a sortie des trioses phosphates du saccharose.
Cette réaction est catalysée par la saccharose-P synthase. Si on isole les chloroplastes et qu’on leur
donne du CO2C14, on n’observe jamais la biosynthese de saccharose. Cependant, si l’on donne du
CO2C14 à des protoplastes, ils fabriquent du saccharose. La biosynthèse du saccharose se fait dans le
cytoplasme. Dans le stroma du chloroplastres les trioses phosphate ald PG et DHAP sont convertis
l’un dans l’autre par isomérisation. Ils sont transportés vers le cytoplasme en échange d’un
phosphate inorganique grâce à des transporteurs spécifiques associés à l’enveloppe du chloroplaste.
Ces transporteurs discriminent préférentiellement le DHAP.
Le DHAP est converti en glucose-1-Phosphate. Ce dernier va être associé à une molécule très riche en
énergie : l’UDP ou Uridine (ou Uritine ???) triphosphate. Ensuite intervention de la phosphate
phosphatase pour donner du saccharose.
IV) synthèse de l’amidon.
Elle est analogue à la voie de biosynthèse du saccharose. Toutefois a partir du glucose-1-Phosphate,
c’est de l’ATP qui est utilisé et non de l’UDP (ou UTP ??)
La photorespiration
I) Introduction
A) Faits expérimentaux
1) Mesure des intensités de photosynthèse apparente
2)
+ ajout
Avec des plantes en C3, plus on augmente l’intensité lumineuse, plus on observe une augmentation
du CO2 produit (ou de l’oxygène absorbé) en fonction de l’intensité lumineuse. Il existe donc chez les
plantes, une forme particulière de respiration dépendante de la lumière : c’est la photorespiration.
3) Conclusion
Dans le cycle de Calvin, nous avions vu que le Ribulose-5-Phosphate est l’accepteur du CO2 et que
1C5 fixe un CO2 (1C1) pour donner deux C6 : c’est la carboxylation du ribulose-1,5-bisphosphate.
Mais l’enzyme qui intervient (la rubisco) est à la fois carboxylase et oxygénase, elle est donc capable
aussi de fixer l’oxygène. Quand elle agit par sa fonction oxygénase, la rubisco fait réagir le ribulose-
1,5-bisphosphate avec l’oxygène. Cette réaction produit un seul APG (C3) et un C2 (Acide
phosphoglycolique)
Il s’agit d’un processus catabolique avec perte de CO2 et absorption d’oxygène, qui consomme de
l’atp et du nadph. La photo respiration diminue l’efficacité de la photosynthèse en dégradant une
partie des sucres formés. La photorespiration dépend du rapport CO2/O2. Elle est supprimée en
présence de forte concentration en CO2. Elle fait intervenir 3 compartiments cellulaires : les
chloroplastes, les peroxysomes, et les mitochondries.
II) Origine du CO2 photorespiratoire.
III) Métabolisme de l’acide phosphogllycolique

La photorespiration se déroule en même temps que la photosynthèse et a la lumière. Elle se traduit
Par une consommation d’o2 et un degagement de CO2. Le cycle se déroule dans 3 compartiments
cellulaires différents : chloroplastes peroxysomes et mitochondries.
1) Dans le chloroplastes : en présence d’oxygène moléculaire, la rubisco catalyse l’oxydation du
ribulose 1,5 biphosphate en phosphoglycérate (C3) et en phosphoglycolate (C2). Le
phosphoclycérate est déphosphorylé en glycolate, et il migre dans les peroxysomes. ceux-ci
sont etroitement accolés au cloroplastre. A ce niveau, le glycolate est oxydé en glyoxylate. Il
y a production de peroxyde d’hydrogene (eau oxygénée = toxique). Ce produit sera
décomponsé par la catalase. Le glyoxylate est alors converti en glycine. La glycine migre dans
les mitchondries.
2) Dans les mitochondries : deux molécules de glycine produisent une molécule de céryne, une
molécule de CO2 et une molécule d’ammoniac NH3. La céryne retourne dans le peroxysome
où elle est convertie en hydroxyde pyruvate.l’hydroxyde pyruvate est réduit en glycérate. Le
glycérate est ensuite phosphorylé soit dans le cytoplasme soit dans le stroma du
chloroplaste.
L’ammoniac produit par la mitochondrie peut être récupérée de deux façons : soit en étant
transformée en glutamate lequel retourne au peroxysome, soit par synthèse de la glutamine dans le
cytoplasme avec formation de deux glutamate. Ce second système est plus efficace que le 1er car la
production d’ammoniac (NH3) par la photorespiration est très importante.
Energie nécessaire à la photorespiration : la photorespiration consomme beaucoup d’énergie. Elle
semble être une reaction inutile. Elle détourne une partie du pouvoir réducteur formé au cours des
réactions lumineuses. Elle n’est pas accompagnée de phosphorylation oxydative (respiration
mitochondriale). Elle est active chez les végétaux en c3 et elle peut inhiber la formation nette de
biomasse de plus de 50%
Stratégies adoptées par quelques plantes pour supprimer la photorespiration.
I) introduction
pour diminuer les effets négatifs de l’activité oxygénase de la rubisco, il faudrait augmenter la
concentration en CO2. Plsuieurs groupes de plantes possèdent des mécansimes spécialisés au niveau
des sites de carboxylation. Ceci diminue l’activité oxygénase et supprime la photorespiration. Les
plantes en C4 ne présentent pas de photorespiration mesurable. Les cellules du mésophile ne
possèdent pas de rubisco. Elles contiennent des enzymes spécialisées insensibles à l’oxygène qui
incorporent le CO2 dans des acides en C4. Ces acides sont transportés vers les cellules du
parenchyme de la gaine périvasculaire qui entoure le faiseau conducteur. Ces cellules possèdent des
mécanismes qui décarboxyle ces acides et liberent le CO2. Elles possèdent également toutes les
enzymes du cycle de calvin des plantes en C3 y compris la rubisco. Le mécanisme d’incorporation du
CO2 des cellules du mésophyle dans des acides est un moyen d’apporter ce cO2 a des concentrations
plus fortes aux cellules de la gaine perivasculaire où la rubisco est localisée.
??? pour la canne a sucre, après 15 secondes de pulse, la radioactivité est retrouvée dans le malate
et dans l’aspartate,( puis dans l’acide 3 phsophoglycérique ?). les exosephosphates peut marquée au
debut de l’incoporation voient leur marquage augmenté après 60 secondes avant de décliner.
Les cellules photosynthétiques des plantes en C4 sont de deux types avec des cloroplaste
différents. Les cloroplastres des cellules du mésophylle sont capables de réaliser une photosynthèse
acyclique avec production d’atp et de NADPH et libération d’oxygène. Elle ne possèdent pas de
rubisco et ne font pas l’incorporation du CO2 en C3. Elles sont riches en phosphoenolpyrubate
carboxylase. Les clhoroplastres de la gaine perivasculaire possede peu de photosystème II et
réalisent la photosynthèse cyclique sans production d’oxygène. Elles contiennent la rubisco qui … ??
ces deux types cellulaires sont voisins et communiquent entre eux par des plasmodesmes.
L’incorporation du CO2 est réalisé dans le cytoplasme des cellules du mésophylle par la PEPC.
L’accepteur du CO2 est le phosphoenol pyruvate. Il est regenéré par un cycle de réaction qui se

déroule succesivement dans les cellules de la gaine vasculaire et dans celle du mésophylle : c’est le
cycle de Hatch et Slack (schéma a savoir) : le CO2 penetre dans les feuilles au niveau des cellules du
mésophylle. Il rentre dans la cellule où il passe sous la forme HCO3-. Il reagit avec le PEP pour donner
l’oxalloacétate. C’est la PEPcarboxylase qui intervient. L’oxaloacétate formé rentre a nouveau dans le
chloroplaste où il est transformé en malate. Le malate sort du cloroplastre, passe par les
plasmodèsme, entre dans le clhoroplastre de la gaine périvasculaire. Dans le chloroplastes, le CO2
porté par le malate est libéré. Il est utilisé par la rubsico pour faire fonctionner le cycle de Calvin. Du
pyruvate est formé à partir du malate. Le pyruvate retourne par les plasodemes dans les
chloroplastes des cellules du mésophylle ou il est transformé en ??
III) les plantes CAM
De nombreuses plantes succulentes adaptés à la sécheresse présente un mécanisme
photosynthétique originale (photosynthese type CAM). Ce métabolisme est proche des palntes en c4
puisquil présente une prémiere incorporation dans un composé organique à 4 carbone./ ce
métabolisme ne se réalise pas dans deux types . ??? la disjonction entre la fixation porimaire du cO2
dans le malate et lincorporation finale via le cycle de calvin se réalise en deux temps. La nuit les
stomates sont ouverts et le CO2 est incorporé dans le malate. celui-ci est mis en réserve dans la
vacuole des cellules. Le jour, les stomates sont fermées. Le malate sort de la vacuole et il est
décarboxylé. Le cO2 libéré est alors incorporé par la rubisco c’est-à-dire le cycle de calvin.
Intérêt biologique des plantes a metabolisme carboxylique
La formation des lgucides a partir du CO2 et de l’eau, suit les mêmes voies biochimiques
chez les plantes en C3, C4 ou cam. Elles utilisent les mêmes longueur d’ondes. La
photosynthèse est fondée sur la fixation du cO2 sur le ribulose biphosphatr et la formation
d’atp et de pouvoir réducteur au niveau des tylakoides. Le metabolisme carboxlique (fication
du CO2 et formation d’acide di carboxylique) intercale en fait la fixation du carbone entre
l’entrée du CO2 et la rubisco ces acides existes aussi chez les plantes en C3 mais il a pris une
place prépondérante chez les plantes en C4. Malgré tout ce syteme consome beaucoup
d’énergie. ???La fixation du CO2 est plus efficace. La photosynthèse d’une plante en C3
s’arrete si la concentration en CO2 est inférieure à 50 ppm. En plante en C4 syntetise jusqu’à
1 ou 2 ppm. La photosynthese en C4 sera donc favorisée dans les régions chaude. Il y aura
formation de glucide même a base pression de CO2 (cas de fermeture des stmates. Il y a
disparition de la compétion CO2 O2 au niveau de la rubisco. La photorespiration est effacée.
La productivité est plus improtante et les plantes fctnne même en eclairement intense (Les
plantes en C3 sont plus efficaces en éclairement faible). La consommation d’eau est plus
faible car elles peuvent syntétiser stomates fermés. Les pertes en eau sont donc limités. Les
plantes en C4 assimillent deux a 3 fois plus de CO2 pour la même quantité deau.
Tous ces avantages expliuquent ds quels régions ces plantes a metaboolisme carboxylique
ont pu se développer. Les plantes Cam sont des plantes de régions très aride a
apprivosinnement en eau difficile ou la secheresse interdit les échanges gazeux. Elles
occupent également des terrains salés. Elles ont peut d’intérêt économique suaf l’ananas.
Les plantes en C4 sont originiaires de régions tropicales ou salés. Ce sont de splantes
herbacéé de grande importance économiqe.
(
Fructose BP
Xylulose P
Xybulose P
Rubisco P
Erythose P
)
1
/
4
100%