Reproduction Pr Barlier 12 pages Métabolisme des glucides

HORMONOLOGIE - REPRODUCTION – Métabolisme des glucides : néoglucogenèse et métabolisme du glycogène
19/03/2015
NAUD Romain L3
CR : AUDOUARD Justine
Hormonologie - Reproduction
Pr Barlier
12 pages
Métabolisme des glucides : néoglucogenèse et métabolisme du glycogène
Les grandes voies métaboliques :
•Glycolyse
•Cycle de Krebs (ou cycle de l'acide citrique)
•Voie des pentoses phosphates
•Néoglucogenèse (synthèse du glucose)
•Glycogène (synthèse et dégradation)
•Biosynthèse et dégradation des AG
•Biosynthèse et dégradation des AA
Aujourd'hui nous allons voir celles qui sont en gras.
A. Première partie : néoglucogenèse
I. Introduction à la néoglucogenèse
a. Généralités
Certains tissus (cerveau, globules rouges, le rein, les muscles, etc.) ont besoin d’être approvisionnés de façon
continue en glucose. Seul le foie est capable d'assurer cette fonction par la mobilisation notamment du
glycogène ainsi que par la néoglucogenèse.
1/12
Plan
A. Néoglucogenèse
I. Introduction à la néoglucogenèse
II. Les réactions de la néoglucogenèse
III. Régulation glycolyse/néoglucogenèse
IV. Les maladies rares dues à un déficit enzymatique
B. Deuxième partie : le métabolisme du glycogène
I. Structure et fonctions du glycogène
II. Synthèse du glycogène : glycogénogenèse
III. Dégradation du glycogène : glycogénolyse
IV. Pathologies enzymatiques : les glycogénoses
V. Régulation réciproque entre synthèse et dégradation du glycogène
C. Conclusion : quelques points clés

HORMONOLOGIE - REPRODUCTION – Métabolisme des glucides : néoglucogenèse et métabolisme du glycogène
La néoglucogenèse est l’ensemble des réactions du métabolisme qui mènent à la synthèse du glucose à partir de
précurseurs non glucidiques (pyruvate, lactate, glycérol et la plupart des acides aminés). Les trois substrats
essentiels du glucose sont l’alanine (obtenu par la dégradation des protéines), le glycérol (obtenu par lyse des
triglycérides) et le lactate (obtenu par des processus anaérobies).
Les réserves du foie sous forme de glycogène sont évaluées à 190g sachant que les besoins journaliers par
individu en glucose sont de 120g pour le cerveau (le plus gros consommateur de glucose), 40g pour le reste de
l’organisme et dans les fluides circulent 20g de glucose à l’état dissout. Les réserves hépatiques ne suffisent pas
pour subvenir aux besoins d’un seul jour. Il faut donc par le biais de l'alimentation subvenir à ces besoins.
b. Biosynthèse du glucose à partir du pyruvate
La voie de la néoglucogenèse convertit le pyruvate en glucose mais ce n’est pas proprement dit la voie inverse
de la glycolyse. En effet, dans la glycolyse certaines étapes sont irréversibles. Il va donc falloir les contourner
par certains systèmes enzymatiques.
Il y a donc différentes étapes qui vont être modifiées par rapport à la glycolyse. Ces deux voies métaboliques
(glycolyse et néoglucogenèse) ont 7 réactions communes tandis 3 réactions se font par des enzymes différentes.
La néoglucogenèse nécessite 3 compartiments intracellulaires : mitochondrial, cytoplasmique et le réticulum
endoplasmique. Ces trois compartiments vont donc être le siège de réactions enzymatiques.
II. Les réactions de la néoglucogenèse
« Je vais insister un peu sur les enzymes car ce sont les enzymes qui sont importantes et qui vont permettre de
contourner les étapes irréversibles de la glycolyse ».
a. Conversion du pyruvate en Phosphoénolpyruvate (PEP)
Dans la glycolyse, la réaction catalysée par la pyruvate kinase est irréversible et ne peut donc pas servir de point
de départ à la néoglucogenèse. Cette réaction est donc substituée par deux autres réactions qui utiliseront un peu
d'énergie (ATP/GTP), ce qui initie le début de la néoglucogenèse et rend possible la conversion du pyruvate en
glucose en le transformant tout d'abord en PEP.
Les enzymes nécessaires pour contourner la pyruvate kinase sont la pyruvate carboxylase qui transforme le
pyruvate en oxaloacétate et la phosphoenolpyruvate carboxykinase (PEP CK) qui le transforme en PEP.
Le pyruvate est exporté dans la mitochondrie, puis est carboxylé par la pyruvate carboxylase au dépend de
l’ATP.
L’oxaloacétate ne peut pas quitter la mitochondrie (il n'a pas de transporteur spécifique) donc on le transforme
en malate qui peut être transporté vers le cytosol (il existe un transporteur spécifique du malate) après
utilisation de la malate déshydrogénase. Puis le retour à l’oxaloacétate par la malate déshydrogénase se fait
dans le cytosol. Enfin, retour au PEP par la PEP CK. Cette PEP CK est réprimée par l’insuline (afin de limiter la
néoglucogenèse et de favoriser la glycolyse).
2/12

HORMONOLOGIE - REPRODUCTION – Métabolisme des glucides : néoglucogenèse et métabolisme du glycogène
b. Transformation du PEP en Fructose-1,6-BiPhosphate (FBP)
La transformation du PEP en FBP se fait grâce à la réversibilité des réactions de la glycolyse.
c. Etapes de conversion du F1,6BP en Glucose
Le FBP est ensuite transformé en Fructose-6-Phosphate (F6P) par la Fructose
1,6-BiPhosphatase.
Ensuite le retour au G6P se fait par une isomérase (commune à la glycolyse). Puis
on a un retour au glucose par la Glucose-6-Phosphatase.
Ces enzymes (différentes entre les deux voies) vont être le siège de régulations très fine par l'insuline et le
glucagon, permettant une balance adaptée entre glycolyse et néoglucogenèse en fonction des situations.
Il y a une certaine importance des trois compartiments (mitochondrie, cytoplasme et RE). En effet, dans la
mitochondrie on a la pyruvate carboxylase, dans la membrane du RE est ancrée la G6Pphosphatase (dans le foie
et le rein uniquement donc absente des muscles et du cerveau) et dans le cytosol le reste des enzymes. Chaque
réaction se fait dans le compartiment contenant l'enzyme nécessaire.
3/12

HORMONOLOGIE - REPRODUCTION – Métabolisme des glucides : néoglucogenèse et métabolisme du glycogène
III. Régulation glycolyse/néoglucogenèse
Chez l’homme le foie métabolise la majeur partie du lactate produit en anaérobie (ce lactate est responsable des
crampes musculaires lorsqu'il est en excès). Ce lactate diffuse dans la circulation sanguine depuis le muscle
vers le foie pour devenir du glucose après une étape intermédiaire en pyruvate. Si le glucose est en excès, il sera
transformé en glycogène.
a. Rapport ATP/AMP
Ces deux étapes se déroulent de façon assez importante dans le cytosol et la plupart des métabolites
intermédiaires sont en communs (7 réactions réversibles). Il va donc y avoir des phénomènes de compétition.
La régulation réciproque des deux voies s'impose et permet de les ajuster en fonction des besoins énergétiques
cellulaires. Les besoins énergétiques conditionnent le sens de la réaction. Ce besoin énergétique s’exprime par
le rapport ATP/AMP.
Si le rapport ATP/AMP est faible, cela signifie que tout l’ATP a été utilisé. On a alors une activation de la
glycolyse (avec activation de la phosphorylation oxydative).
Si le rapport ATP/AMP est élevé, cela signifie que l'ATP et ses précurseurs sont en quantité suffisante. On a
alors un ralentissement de la glycolyse, l'excès de pyruvate transformé en glucose suite à une activation de la
néoglucogenèse (pour créer du glycogène et stocker les réserves).
b. Régulation hormonale
Deux hormones principales : l’insuline et le glucagon
Elles sont importantes, s’il y a un dérèglement de l'une d'entre elle apparaît une pathologie comme le diabète.
L’insuline est sécrétée lorsque le taux de glucose dans le sang est élevé. Elle va être sécrétée lors d’apport en
glucose importants. Elle a plusieurs actions principales :
•Augmente l’entrée de glucose dans les tissus périphériques
•Active la glycolyse et la synthèse de glycogène
•Inhibe la néoglucogenèse
Le glucagon est sécrété lorsque le taux de glucose dans le sang est faible :
•Active la dégradation de glycogène
•Active la néoglucogenèse dans le foie
•Avec ces deux phénomènes : libération de glucose du foie dans le sang
IV. Les maladies rares dues à un déficit enzymatique
Les maladies rares sont en rapport avec les déficits enzymatiques. L’ensemble de ces pathologies sont recensées
sur orphanet. Lorsqu'une enzyme est absente ou en quantité insuffisante dans l'organisme, cela va entraîner des
problèmes au niveau de la physiologie de la régulation de la glycémie.
4/12

HORMONOLOGIE - REPRODUCTION – Métabolisme des glucides : néoglucogenèse et métabolisme du glycogène
a. Exemple du déficit en PEP CK
Elle a une hérédité mitochondriale (la PEPCK se trouve dans la mitochondrie), sa prévalence est très faible (< 1
/ 1000000).
Le déficit de cette enzyme entraîne un trouble de la néoglucogenèse et se manifeste par des hypoglycémies, une
détérioration neurologique progressive (car il n’y a plus de glucose dans l’organisme notamment à l’état de
jeûne et ça atteint le cerveau, principal consommateur de glucose), une insuffisance hépatique sévère, un retard
de développement et le syndrome de Fanconi.
b. Exemple du déficit en Glucose-6-Phosphatase
Il existe d’autres déficits comme la glycogénose du à un déficit G6Phosphatase.
Elle entraîne une intolérance au jeûne (on va pas pouvoir produire le glucose). On peut avoir un retard de
croissance, une hépatomégalie (par accumulation de glycogène). Elle peut se manifester dès la naissance par
une hépatomégalie et des symptômes d’hypoglycémie induite par le jeûne.
B. Deuxième partie : le métabolisme du glycogène
I. Structure et fonctions du glycogène
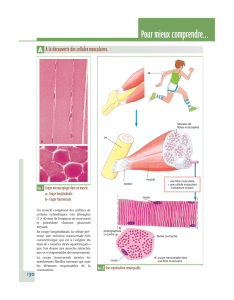
En histologie, on observe des granules de glycogènes qui peuvent s’accumuler dans le foie.
C’est un poly-glucose qui forme une mise en réserve de glucose rapidement mobilisable. Il représente la
majeure partie des sucres stockés chez les mammifères et est présent dans le foie et les muscles, au niveau du
cytosol. C’est l’analogue de l’amidon chez les végétaux.
Le glycogène est un polymère de glucoses (uniquement), en alpha-D-glucopyranose avec des liaisons de type
α-1,4 ainsi que des branchements reliées à la molécule principale par des liaisons glycosidiques de type α-1,6.
C'est une molécule relativement très branchée.
5/12
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%