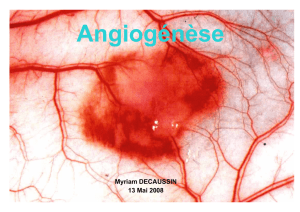
Effets extravasculaires neurologiques des VEGF

Effets extravasculaires neurologiques
des VEGF
Bernard Lévy
Institut des vaisseaux et du sang, Hôpital
Lariboisière, Paris, France
<bernard.levy@inserm.fr>
3
VEGF Actu
N° 34 • Mars 2014
Les différentes isoformes du VEGF ont
des effets qui dépassent leurs rôles
dans le contrôle de la perméabilité vascu-
laire et de l’angiogenèse. Les VEGF et
leurs récepteurs (VEGFR) interviennent
également dans le développement du
système nerveux et sont actuellement
considérés comme des acteurs majeurs
de la neurogenèse. Il y a 20 ans, le groupe
de Werner Risau avait, le premier, rapporté
que les cellules embryonnaires cérébra-
les expriment fortement le VEGFA le long
des parois ventriculaires [1] ; Eli Keshet
montrait, quant à lui, que l’expression de
VEGFA est régulée par l’hypoxie dans le
glioblastome [2] ; enfin Connie Cepko
observait, peu après, que le VEGFR2 (Flk-
1, KDR) est exprimé à la surface des cellu-
les nerveuses progénitrices neuronales
dans la rétine de la souris, ce récepteur
jouant un rôle indispensable dans la neuro-
genèse rétinienne [3]. Depuis ces travaux
fondateurs, de nombreuses équipes
essaient de comprendre comment la
famille des VEGF et leurs récepteurs parti-
cipent au développement, puis au main-
tien et à la réparation des cellules nerveu-
ses. Tous les fondamentalistes travaillant
sur les VEGF soulignent que cette famille
de molécules a des effets pléiotropiques
puissants sur le système nerveux. Le trai-
tement de certaines maladies neurologi-
ques dégénératives par des facteurs de
croissance vasculaire est très sérieuse-
ment envisagé. Nous souhaitons vous
présenter plus spécialement les effets du
VEGF sur la plasticité neuronale et les
fonctions cognitives, peu connus dans le
domaine de l’oncologie. Il n’est pas impos-
sible que certains effets secondaires des
anti-angiogéniques soient en relation,
directe ou indirecte, avec leurs effets sur
le système nerveux central [4]. Dans le
numéro 22 de VEGF Actu (mars 2011),
Sophie Taillibert avait traité des atteintes
cognitives méconnues liées aux traite-
ments anti-angiogéniques. Elle soulignait
le fait que « le plus souvent subtile et dura-
ble, cette atteinte cognitive qualifiée de
“modérée” par les médecins, affecte les
activités quotidiennes et doit être recher-
chée. »
On considère qu’il n’y a pas d’angioge-
nèse notable dans le cerveau adulte dans
les conditions normales ; cependant,
comme dans tous les organes, le turn over
naturel des cellules vasculaires nécessite
un renouvellement très lent et quantitati-
vement limité de ces cellules. De plus,
une augmentation durable de l’activité
métabolique cérébrale s’accompagne
d’une augmentation de la densité micro-
vasculaire locale dans la région du cerveau
concernée par l’augmentation d’activité.
Le VEGF, en raison du contrôle exercé sur
lui par l’hypoxie et l’hypoglycémie loca-
les, est le facteur essentiel qui permet
l’adaptation de la densité microvascu-
laire locale à la consommation tissulaire
d’oxygène [5].
L’enrichissement environnemental permet
d’étudier l’influence du contexte exté-
rieur sur le cerveau dont les centres de
perception sont affectés par les stimula-
tions sensorielles multiples, y compris les
interactions sociales. Exposé à un envi-
ronnement plus riche et stimulant, le
nombre de synapses augmente dans le
cerveau. Cet effet est plus important durant
la phase de développement du système
nerveux, mais persiste néanmoins à un
degré moindre pendant l’âge adulte. L’aug-
mentation du nombre et de la complexité
des synapses génère également une acti-
vité électrique plus importante, ce qui
entraîne une augmentation de la taille et
du nombre des cellules gliales. Le cortex
cérébral est alors plus épais et on peut
observer une neurogenèse significative,
du moins chez les rongeurs. Les travaux
expérimentaux montrent qu’un environ-
nement plus stimulant peut aider dans le
traitement et la restauration d’une grande
variété de maladies cérébrales telles la
sénilité et la maladie d’Alzheimer. Les
recherches effectuées chez l’homme suggè-
rent qu’un déficit de stimulation retarde
et contrarie le développement cognitif du
nouveau-né et de l’enfant. Chez l’adulte,
les individus bénéficiant des plus hauts
niveaux d’éducation, supposés avoir une
stimulation cognitive élevée, ont une plus
grande résilience contre les effets de l’âge
et des démences dégénératives. L’élevage
des souris dans un environnement « enri-
chi », augmente la densité microvascu-
laire dans l’hippocampe ; chez le rat, on
a montré une corrélation entre la
complexité des stimuli visuels, la densité
microvasculaire et l’expression du VEGF
dans le cortex visuel [6].
Différentes méthodes ont été utilisées
pour étudier l’importance de la signalisa-
tion continue des récepteurs du VEGF sur
la vascularisation cérébrale. Chez l’adulte,
il semble que le réseau vasculaire dépende
peu ou pas de la voie des VEGF en compa-
raison avec d’autres organes [7, 8]. Une
exception notable concerne les plexus
choroïdes. Ces réseaux vasculaires parti-
culiers contiennent de nombreux capillai-
res dont l’endothélium est très fenêtré. Le
liquide cérébrospinal est sécrété, à partir
du sang, à travers ces fenêtres endothé-
liales. Lors de l’inhibition de la voie du
VEGF, la vascularisation des plexus choroï-
des régresse et la perméabilité endothé-
liale diminue, de même que la sécrétion
de liquide cérébrospinal [9]. On sait en
effet que la taille, le nombre et la perméa-
Brève
Figure 1. Expression
endogène de VEGF
dans le cerveau
de la souris adulte
(marquée en bleu par
LacZ). Les astrocytes
sont colorés en brun
(GFAP). CA1 :
l’une des parties de
l’hippocampe, DG :
gyrus dentelé,
SVZ : zone supra-
ventriculaire, RMS :
courant de migration
rostrale, OB : bulbe
olfactif. D’après [15].
VEGF-34 maquette2:VEGF-maq-18-d4 15/04/14 10:18 Page3
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

4VEGF Actu
N° 34 • Mars 2014
Brève
bilité des fenestrations capillaires sont
très dépendants des concentrations loca-
les de VEGF [8].
L’importance du VEGF dans le cerveau
est suggérée par l’évaluation quantitative
de son expression dans les différentes
zones cérébrales [10] (figure 1). Comme
on peut s’y attendre, l’expression la plus
importante est retrouvée, chez l’adulte,
au niveau des plexus choroïdes. Dans le
parenchyme cérébral de l’adulte, les astro-
cytes sont la première source de VEGF.
On sait que les astrocytes sont les cellu-
les cérébrales qui répondent le mieux à
une hypoxie locale par une augmenta-
tion de l’expression de HIF1 [11]. Il est
logique de supposer que les astrocytes
sont les « senseurs » cérébraux les plus
sensibles à l’hypoxie et que l’expression
de VEGF, cible directe de HIF1, reflète
cette sensibilité à la pression partielle
locale d’oxygène.
Dans certaines régions du cerveau, le
VEGF est préférentiellement exprimé par
des cellules autres que les astrocytes :
dans l’hippocampe, les neurones pyra-
midaux CA1 expriment fortement le VEGF ;
dans le cortex, le VEGF est exprimé par
certaines cellules pyramidales ; dans le
cervelet, par les cellules de Purkinje. Il
faut souligner que dans toutes ces locali-
sations, le réseau vasculaire adulte est
quiescent, suggérant que l’expression
constitutive de VEGF correspond à des
fonctions déconnectées de ses fonctions
angiogéniques.
VEGF et plasticité
synaptique, activité
neuronale, apprentissage
et comportement
La plasticité synaptique se définit comme
la capacité des synapses à se modifier en
fonction d’une sollicitation intensive ou,
au contraire, faible ou absente. La plasti-
cité synaptique est probablement un
élément essentiel des processus d’ap-
prentissage et de mémorisation. Les méca-
nismes moléculaires qui commandent la
plasticité synaptique ne sont pas bien
connus. Ils impliquent certainement les
neurotransmetteurs produits et libérés
dans l’espace synaptique. La quantité de
neurotransmetteur en jeu, la réponse post-
synaptique à ces neurotransmetteurs et
les modifications de nombre et de la
densité des synapses sont certainement
des éléments importants qui condition-
nent leur plasticité. Il est probable que le
VEGF est impliqué dans ces mécanismes
dynamiques. L’application de VEGF sur
des tranches d’hippocampe étudiées in
vitro diminue l’amplitude des potentiels
évoqués dans tout l’hippocampe et inhibe
les décharges électriques spontanées dans
des modèles de rat épileptique [12]. A
l’inverse, dans des cultures de cellules de
l’hippocampe, HIF1 et VEGF augmentent
l’activité électrique et chimique des synap-
ses [13]. Les études les plus convaincan-
tes et les plus proches des conditions in
vivo suggèrent que le VEGF augmente
considérablement la plasticité neuronale
alors que l’inhibition de la voie du VEGF
bloque cette plasticité. Les neurones,
astrocytes et vaisseaux sanguins forment
des unités neuro-vasculaires dans lesquel-
les le vaisseau joue un rôle d’ajustement
important. Dans un modèle animal très
sophistiqué qui permet d’explorer sépa-
rément l’angiogenèse, la neurogenèse et
la mémoire, E. Keshet [14] a montré que
la surexpression du VEGF augmente ces
3 paramètres alors que le blocage de la
voie du VEGF induit une détérioration de
la mémoire sans modifier la perfusion de
l’hippocampe ni la neurogenèse.
De nombreux travaux expérimentaux
démontrent que l’administration ou la
surexpression de VEGF augmentent les
capacités d’apprentissage et la mémoire
par des mécanismes localisés dans l’hip-
pocampe [15]. Cela est à rapprocher du
fait que les astrocytes de l’hippocampe
sécrètent du VEGF chez l’animal soumis
à un environnement « enrichi », à un exer-
cice musculaire, à des sollicitations mémo-
rielles ou à une stimulation électrique de
l’hippocampe. Il semble donc clair que,
dans les modèles animaux, le VEGF parti-
S’il existe bien une caractéristique commune
aux conséquences du ciblage de l’angio-
genèse en clinique, c’est que les effets des
traitements sont multiples avec parfois des
conséquences semblant diamétralement
opposées. Il en est ainsi également de l’in-
teraction entre angiogenèse cérébrale,
cancers et traitements anti-angiogéniques.
L’analyse des conséquences éventuelles
sur les fonctions cognitives (positives ou
négatives) de l’utilisation d’anti-angiogéni-
ques dans le contexte général de la prise
en charge d’une pathologie maligne est
complexe du fait : (1) de l’existence parfois
de lésions cérébrales ou d’autres traite-
ments entraînant eux-mêmes des troubles
cognitifs ; (2) du terrain clinique sous-jacent
de la maladie qui peut lui-même être un
facteur prédisposant à des troubles cogni-
tifs ; et (3) de l’utilisation de médicaments
anti-angiogéniques ayant des spectres d’ac-
tion différents, plus ou moins spécifiques
de l’angiogenèse. J’illustrerai mes propos
par quelques exemples récents de la litté-
rature.
• La prise en charge de lésions cérébrales
malignes primitives ou secondaires en radio-
thérapie peut se compliquer de radioné-
crose entraînant des troubles neurologi-
ques importants pouvant engager le
pronostic fonctionnel, voire vital, du patient
traité. Une analyse récente de la littérature
montre l’intérêt du bevacizumab dans la
prise en charge de ces complications avec
une amélioration radiologique et clinique
[1]. Agir sur le VEGF est une des possibili-
tés de réduire la toxicité cérébrale de la
radiothérapie.
• Deux études récentes ont rapporté l’effet
d’un traitement par bevacizumab dans la
prise en charge du glioblastome. S’il n’y a
pas d’amélioration de la survie globale,
une des deux études conclut à un contrôle
plus long de la maladie, de 3 à 4 mois. Toute-
fois, si l’on se focalise sur l’évaluation des
fonctions cognitives des patients inclus,
une des études retrouve un maintien des
fonctions sous traitement, et l’autre une
altération… sans que nous ayons une expli-
cation rationnelle de cette différence fonda-
mentale qui, à elle seule, peut être un argu-
ment pour prescrire ou non ce traitement
lorsque la survie semble inchangée [2]…
• Le sorafenib est utilisé dans le traitement
du carcinome hépato-cellulaire survenant
très fréquemment chez des patients porteurs
de cirrhose hépatique à risque d’encépha-
lopathie. Des collègues italiens ont récem-
ment rapporté que le sorafenib pouvait
être un facteur déclenchant une encépha-
lopathie métabolique chez ces patients
[3]. Plus étonnant, l’encéphalopathie semble
ne pas être sensible au traitement classi-
que mais est totalement réversible à l’arrêt
du sorafenib, laissant supposer qu’une action
neuronale directe de celui-ci puisse déclen-
cher l’apparition de l’encéphalopathie
métabolique chez certains patients…
La complexité des situations cliniques rend
difficile l’interprétation de l’impact cogni-
tif de ces traitements, mais n’en demeure
pas moins un enjeu important dans la prise
en charge au long cours de nos patients,
nécessitant une analyse rigoureuse des
effets et du terrain dans les études menées.
Références
1. Tye K, et al. J Neurooncol 2014 : Feb 7 [Epub
ahead of print].
2. Fine HA. N Engl J Med 2014 ; 370 : 764-5.
3. Brandi G, et al. Liver Int 2013 ; 33 : 488-
93.
Angiogenèse cancer et cerveau en pratique
clinique : le yin et le yang ?
Joseph Gligorov
VEGF-34 maquette2:VEGF-maq-18-d4 15/04/14 10:18 Page4
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.

5
VEGF Actu
N° 34 • Mars 2014
Revue
cipe à l’amélioration des activités cogni-
tives.
Dans l’hippocampe, le VEGF agit à la
fois sur la neurogenèse, l’angiogenèse et
les fonctions cognitives. L’amélioration
des performances cognitives est obser-
vée très tôt après l’augmentation de l’ex-
pression du VEGF dans l’hippocampe,
bien avant l’apparition de toute neuroge-
nèse, suggérant que ces deux effets
(neurogenèse et amélioration des perfor-
mances) ne sont pas liés et que les effets
vasculaires du VEGF pourraient être en
première ligne.
Conclusion
De toute évidence, les effets du VEGF
sur le système nerveux et sur son activité
sont complexes mais indiscutables. Dans
la plupart des cas, on a du mal à savoir si
ces effets sont directs, sur les récepteurs
neuronaux du VEGF, ou indirects par l’in-
termédiaire des cellules gliales et endo-
théliales qui sécréteraient des facteurs
agissant sur les neurones voisins. Reste à
savoir si les différentes classes de médi-
caments anti-angiogéniques ont des effets
secondaires neurologiques en rapport
avec ce qui précède. De manière plus
prospective, on peut imaginer que, un
jour, les maladies nerveuses dégénérati-
ves pourront bénéficier de traitements
pro-angiogéniques.
Liens d’intérêts : Les auteurs déclarent
ne pas avoir de lien d’intérêt en rapport
avec cet article.
Références
1. Breier G, et al. Development 1992 ; 114 : 521-
32.
2. Shweiki D, et al. Nature 1992 ; 359 : 843-5.
3. Yang K, et al. J Neurosci 1996 ; 16 : 6089-99.
4. Thomas JL, et al. Cell Mol Life Sci 2013 ; 70 :
1673-4.
5. Ueno S, et al. J Cell Physiol 2008 ; 217 : 13-22.
6. Bengoetxea H, et al. Cereb Cortex 2008 ; 18 :
1630-9.
7. Tang K, et al. J Appl Physiol 2004 ; 97 : 1559-66.
8. May D, et al. PLoS One 2011 ; 6 : e21478.
9. Kamba T, et al. Am J Physiol Heart Circ Physiol
2006 ; 290 : H560-H576.
10. Maharaj AS, et al. Am J Pathol 2006 ; 168 : 639-
48.
11. Weidemann A, et al. J Clin Invest 2009 ; 119 :
3373-83.
12. McCloskey DP, et al. J Neurosci 2005 ; 25 :
8889-97.
13. Huang YF, et al. J Neurosci 2010 ; 30 : 6080-93.
14. Licht T, et al. Proc Natl Acad Sci USA 2011 ;
108 : 5081-6.
15. Licht T, et al. Cell Mol Life Sci 2013 ; 70 : 1727-
37.
La lymphangiogenèse
Bases physiologiques et approches
physiopathologiques
Joseph Gligorov1, Bernard Lévy2
Service d’oncologie médicale, Institut universitaire de cancérologie, APHP Tenon, Paris,
France
<joseph.gligorov@tnn.aphp.fr>
2. Institut des vaisseaux et du sang, Hôpital Lariboisière, Paris, France
<bernard.levy@inserm.fr>
Réseau lymphatique
et lymphangiogenèse
Le rôle fonctionnel du réseau lymphati-
que est capital dans la régulation des volu-
mes extracellulaires, le déclenchement
des réponses immunitaires et dans la régu-
lation du métabolisme des lipides. Une
anomalie lymphatique, génétique ou trau-
matique, peut donc entraîner des œdèmes,
une atteinte des défenses immunitaires
et/ou une accumulation des dépôts de
graisses, le plus souvent sous-cutanées.
L’endothélium capillaire systémique est
en principe imperméable aux protéines
de grande taille (> 70 kD) ; en réalité,
une fuite minime de protéines se produit
et entraînerait au cours du temps, si elle
n’était pas compensée, une accumula-
tion de protéines dans le milieu extravas-
culaire et un œdème oncotique. Dans
les conditions normales, la quasi-totalité
des macromolécules qui sortent des
capillaires sanguins est réabsorbée par le
réseau lymphatique.
Le réseau lymphatique est parcouru par
un courant unidirectionnel à très faible
débit (le débit lymphatique total est de
l’ordre de 1 L/24 h). La lymphe se forme
dans des capillaires lymphatiques borgnes
(figure 1) qui se réunissent en canaux
collecteurs drainés par des ganglions.
Les capillaires lymphatiques ont une paroi
plus fine et une lumière plus grande que
les capillaires sanguins ; ils ne contien-
nent pas de péricytes et ne comportent
pas de membrane basale. Des filaments
d’ancrage lient les capillaires lymphati-
ques à la matrice extracellulaire et assu-
rent leur stabilité anatomique. Une partie
des fluides et toutes les macromolécules
qui sont sorties des capillaires sanguins
entrent dans le capillaire lymphatique
borgne à travers les interstices existant
entre les cellules endothéliales qui jouent
le rôle de micro-valves. Dans les canaux
collecteurs, on trouve des cellules endo-
théliales jointives, donc moins perméa-
bles, une membrane basale, des péricy-
tes, des cellules musculaires lisses et de
véritables valves anatomiques qui empê-
chent le reflux lymphatique.
Les vaisseaux lymphatiques transportent
également les leucocytes sortis des capillai-
res sanguins par extravasation, des
Membrane basale Membrane basale
Péricytes
Péricytes
SMC
Fluide
et macro-
molécules
Filaments
d’ancrage
Valve lymphatique
Microvalve
Capillaire sanguin Capillaire lymphatique Vaisseaux lymphatiques
Figure 1. Structure des vaisseaux lymphatiques. Membrane basale, capillaire sanguin,
péricytes, liquide et macromolécules, capillaire lymphatique, filaments d’ancrage,
microvalves, SMC = cellules musculaires lisses, valve lymphatique, vaisseau lymphati-
que collecteur.
VEGF-34 maquette2:VEGF-maq-18-d4 15/04/14 10:18 Page5
Copyright © 2017 John Libbey Eurotext. Téléchargé par un robot venant de 88.99.165.207 le 25/05/2017.
1
/
3
100%