Sujet de rattrapage - Durée : 2 heures Exercice 1

SVT-Tronc commun : Stabilité, Variabilité des génomes, Evolution
Sujet de rattrapage -Durée : 2 heures
Exercice 1 :
Les levures sont des champignons unicellulaires. Lorsque les conditions sont
favorables, elles se reproduisent par bourgeonnement: toutes les 90 minutes
environ, une nouvelle cellule se forme par mitose en périphérie de la cellule
mère, puis se détache.
Dans des conditions particulières, deux levures peuvent fusionner (étape X du
schéma). La cellule ainsi formée peut bourgeonner àson tour. Ces levures C ne
fusionnent jamais, mais si les ressources nutritives du milieu s’épuisent, elles
subissent deux divisions successives (étape Y) aboutissant à la formation de
quatre cellules génétiquement différentes contenues dans une enveloppe. Dans
des conditions favorables, chacune de ces cellules peut former d’autres levures
par bourgeonnement.
a. (raisonner) En indiquant votre raisonnement, nommez les
étapes X et Y et déterminez le nombre de chromosomes contenus
dans les cellules A, B et C, sachant que le nombre haploïde de
chromosomesest égal à 17 chez la levure.
b. (utiliser ses connaissances, comparer) Quelle(s) particularité(s) de ce cycle le différencie des cycles vus
en cours ?
c. (restituer des connaissances) Illustrez par des schémas les mécanismes chromosomiques mis en jeu au
cours des étapes X et Y du doc.ci-dessus, en limitant le nombre de chromosomes à n=3 ou 2n=6
Exercice 2 :
Un modèle possible de l'histoire évolutive des gènes qui codent la
lactico-déshydrogénase (notée LHD), enzyme présente chez tous
les êtres vivants, est représenté dans le document
de référence.
(extraire des information d’un document pour argumenter sa réponse)
Montrez que les informations apportées par le document
(1a et 1b) valident le modèle proposé de l'histoire
évolutive des gènes de la LDH
Document
La lactico-déshydrogénase (LDH) est une enzyme constituée par
l'association de quatre chaînes polypeptidiques qui peuvent être
identiques ou non. Chez l'Homme il existe trois types de chaînes (LDH-
A, LDH-B et LDH-C).
1a) Localisation des gènes de la LDH sur les chromosomes de
l'Homme :
1b) Séquences partielles alignées d'acides aminés des
chaînes polypeptidiques LDH-A, LDH-B et LDH-C
humaines:
N.B. : dans les séquences de LDH-B et LDH-C, un tiret indique un
acide aminé identique à celui présent dans la séquence de LDH-A,
laquelle est prise arbitrairement comme référence.
Document de
référence :
Modèle proposé pour
illustrer l'histoire
évolutive qui a
conduit aux gènes
qui codent la lactico-
déshydrogénase
chez l'Homme.

Exercice 3 :
On s'intéresse ici à l'origine d'une maladie génétique, la mucoviscidose, et à son maintien dans les populations
humaines.
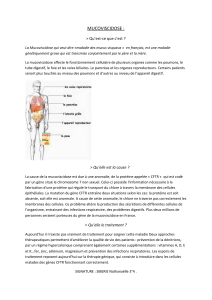
La mucoviscidose est une maladie monogénique (dysfonctionnement d’un seul gène) causée par l’obstruction des bronches et
bronchioles pulmonaires par un mucus anormalement épais, visqueux et abondant.
Les symptômes de la maladie sont des troubles respiratoires et digestifs très graves : l’encombrement par le mucus diminue
fortement les capacités respiratoires du sujet atteint et provoque fréquemment des infections pulmonaires répétitives.
Il y a 20 ans, les malades n’atteignaient que très rarement l’âge adulte, aujourd’hui leur espérance de vie a beaucoup
progressé (25 à 30 ans en moyenne)
DOCUMENT 1:
En 1989, le gène de la mucoviscidose à été découvert par une équipe des universités de Toronto (Canada) et du
Michigan (U.S.A).
La protéine de 1480 acides aminés codée par ce gène est appelée CFTR, c’est une protéine membranaire qui permet les
échanges d’ions Chlorure (Cl-) à travers la membrane plasmique : chez les sujets normaux, l’activité de cette protéine forme
des canaux qui rendent la cellule perméable et permettent le transport des ions Cl-. Chez les individus malades, ces canaux
sont bloqués par une modification de structure de la CFTR, ou sont reconnus comme anormaux et détruits. La conséquence
est la même dans les deux cas : les canaux aux ions chlorures sont inefficaces.
Comme la sortie du Cl-est normalement accompagnée d'une sortie d'eau, les cellules de l’épithélium pulmonaire du malade
sont quasi-imperméables à l'eau, d'où des sécrétions bronchiques anormalement épaisses, cause d'encombrement et
d'insuffisance respiratoire chronique.
Cette maladie est récessive : un seul allèle fonctionnel est suffisant pour assurer la perméabilité nécessaire à une sécrétion
normale du mucus. Cependant dans la population française, 1 personne sur 20 est porteuse d’un allèle muté.
DOCUMENT 2 : tableau du code génétique:
La séquence normale du gène (brin codant) de la
mucoviscidose est la suivante :
..AAA GAA AAT ATC ATC TTT GGT GTT TCC TAT...
Ce gène code pour la synthèse d'une protéine efficace de
1480 acides aminés, dont la séquence partielle est:
..LYS-GLU-ASN-ISO-ISO-PHÉ-GLY-VAL-SER-TYR...
(503)... ...(512)
La version allèlique anormale la plus fréquente (responsable
de 70% des cas de mucoviscidose) présente, pour la même
portion du gène, la séquence suivante:
..AAA GAA AAT ATC ATT GGT GTT TCC TAT...
DOCUMENT 3 :
Salmonella typhi est la bactérie responsable de
la typhoïde. Cette maladie infectieuse se traduit par une
inflammation grave du tube digestif, souvent mortelle
jusqu’à la découverte d’un traitement antibiotique il y a
une cinquantaine d’année.
Des chercheurs ont récemment réussi à intégrer le gène
humain de la protéine CFRT dans le génome de la souris :
on obtient ainsi des souris transgéniques.
Ils ont alors étudié la pénétration de la bactérie dans
les cellules intestinales de trois lots de souris
transgéniques :
(pratiquer un raisonnement scientifique) A l’aide de ces documents, expliquez précisément l’origine moléculaire de
cette maladie et proposez une hypothèse expliquant le maintien de l’allèle muté à une fréquence
relativement élevée.

Exercice 1 : sur 6pts
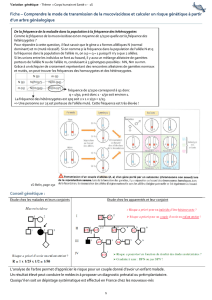
L’étape Xmontre la fusion de deux cellules et de leurs noyaux : cette étape est la fécondation (la
caryogamie) qui forme une cellule œuf diploïde, soit 2n = 34 chromosomes pour la cellule C
L’étape Ymontre une succession de deux divisions : cette étape est la méiose qui forme des spores
haploïdes, soit n=17 pour les cellules A et B.
0.5+0.5
0.5
0.5+0.5
0.5
Dans ce cycle, contrairement aux cycles vu en cours :
- la reproduction conforme par mitose concerne autant la phase haploïde que la phase
diploïde,
-il n’y a pas une phase «dominante » sur l’autre,
-ce sont les conditions du milieu qui déterminent une reproduction sexuée ou une
reproduction conforme.
0.5
0.5
Schéma chromosomique des deux divisions de la méiose : (2n=6 n=3)
Schéma chromosomique de la fécondation : n=3 + n=3 2n=6
Soin des schémas, légendes, titres
1
0.5
0.5
Exercice 2 : Sur 4
La notion de gène ancestral est validée par les ressemblances de séquence constatées entre les 3 protéines au
document b :
14 acides aminés sur les 26 étudiés, soit plus de 50% sont identiques pour les 3 chaînes A B et C
0.75
0.25
L’existence de deux duplications successives
est validée par l’existence de 3 gènes au document a: ces gènes sont situés à des locus différents des
chromosomes 11 (pour A et C) et 12 (pour LDH-B)
0.5
0.5
Les mutations successives peuvent être mises en évidence par les différences entre les séquences peptidiques
puisque la séquence en acide aminée est déterminée par la séquence en nucléotide de l’ADN 0.75
0.25
On constate seulement 7 différences entre LDH-A et LDH-C, 9 différences entre LDH-A et LDH-B et 12
différences entre LDH-B et LDH-C,
donc LDH-A et C sont les plus «proches » génétiquement, ce qui valide la chronologie des duplications
successives (gène ancestral commun àLDH-A et C plus récent que celui partagé avec LDH-B, sachant que
plus la séparation est ancienne et plus les gènes divergent par mutation et donc moins les protéines se
ressemblent.)
0.75
0.25
Exercice 3: sur 6 pts
Le document 1 permet d’expliquer les symptômes de la mucoviscidose par l’altération d’une
protéine (CFTR) dont la structure est modifiée, ce qui la rend non fonctionnelle.
0.5
Le document 2permet d’identifier la mutation responsable de l’allèle anormal :
il s’agit d’une délétion de 3 nucléotides
la séquence peptidique obtenue est anormale car il manque l’acide aminé 508 (PHE):
...LYS-GLU-ASN-ISO-ISO-GLY-VAL-SER-TYR...
la disparition d’un acide aminé (au niveau du site actif) suffit ici à rendre la protéine inefficace.
0.25
0.5+0.25
0.5
La maladie est récessive donc les malades sont homozygotes récessifs, (les homozygotes dominants
et les hétérozygotes sont sains).
0.5
Dans le document 3, on constate que l’allèle muté confère une protection (une résistance) vis-à-vis
de la bactérie Salmonella typhi.
à l’état homozygote ou hétérozygote (lots 2 et 3)
0.5
0.5
Raisonnement :
La mort précoce des malades (empêchant leur reproduction) est une contrainte sélective négative
qui aurait du limiter la fréquence de cet allèle défavorable dans la population.
Or sa fréquence est de l’ordre de 5% : le maintien de cet allèle muté s’explique grâce à une autre
composante de l’environnement, une maladie bactérienne, la typhoïde, qui exerce sur l’allèle muté
une pression sélective positive : elle avantage l’allèle muté.
Il y a ici un mécanisme de sélection naturelle c'est-à-dire un avantage sélectif pour les
hétérozygotes, non atteints par la mucoviscidose et peu touchés par la typhoïde.
Conclusion : puisque l’allèle muté est à la fois défavorable (à l’état homozygote) et favorable (à
l’état hétérozygote), il se crée un équilibre qui maintient la fréquence de cet allèle à environ 1/20.
0.5
0.5
0.5
0.5
0.5
1
/
3
100%