Télécharger le fichier - Fichier

ENZYMOLOGIE GENERALE
Professeur Brigitte LEININGER-MULLER
Faculté de Pharmacie
5, rue A. Lebrun
54000 NANCY
Faculté de Médecine
INSERM U954
Bâtiment C, 2ème étage
PACES 2010-2011

INTRODUCTION
1. NOTIONS GÉNÉRALES
1.1. Catalyse enzymatique
1.2. Définitions
1.3. Localisation et variations physiopathologiques
1.4. Isoenzymes
1.5. Proenzymes ou zymogènes
1.6. Enzymes inductibles
2. STRUCTURE ET SPÉCIFICITÉ DES ENZYMES
2.1. Apoenzyme et coenzyme
2.2. Nécessités auxquelles doit répondre la structure des enzymes
2.3. Site actif
2.3.1. Définition – nature - fonctionnement
2.3.2. Conformation du site actif
2.4. Spécificité des réactions enzymatiques

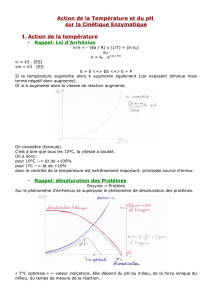
3. CINÉTIQUE ENZYMATIQUE
3.1. Notions de cinétique des réactions
3.1.1. Vitesse de réaction – Vitesse instantanée
3.1.2. Différentes phases de la réaction enzymatique
3.2. Principe de la mesure des vitesses des réactions
enzymatiques – Unités
3.3. Cinétique michaelienne
3.3.1. Variation de la vitesse en fonction du temps
3.3.2. Variation de la vitesse en fonction de la quantité
d’enzyme
3.3.3. Effet de la concentration de substrat - Vitesse initiale
3.3.4. Equation de Michaelis et Menten - Hyperbole de
Michaelis-Menten
3.3.5. Détermination de Km
- Représentation de Lineweaver et Burk
- Représentation de Eadie - Hofstee

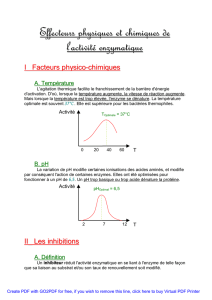
3.4. Effecteurs chimiques de la réaction enzymatique
3.4.1. Activateurs
3.4.2. Inhibiteurs
3.4.2.1. Inhibiteurs compétitifs
3.4.2.2. Inhibiteurs non compétitifs
3.4.2.3. Inhibiteurs incompétitifs
3.4.3. Effecteurs allostériques
4. CLASSIFICATION DES ENZYMES. INTÉRÊT EN BIOLOGIE
4.1. Nomenclature
4.2. Oxydo-réductases
4.3. Transférases
4.4. Hydrolases
4.5. Lyases
4.6. Isomérases
4.7. Ligases

composé qui n’accélère que les réactions thermodynamiquement
possibles, i.e :
celles qui se traduisent par une diminution d’enthalpie libre.
Le catalyseur diminue l’énergie d’activation nécessaire à la réaction.
Un catalyseur
se retrouve à la fin de la réaction tel qu’il était avant la réaction
agit à des concentrations très faibles en comparaison avec les
quantités des composés en réaction
spécifique d’une réaction donnée, c’est à dire qu’il n’active
que cette réaction
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
1
/
54
100%