les enzymes - cloudfront.net

LES
E
NZYMES
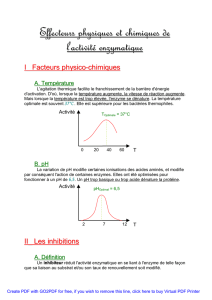
!"#$$%&#!
"'#(#)* !
+ #,&#" , # $ &#-./
0,1&#" #0###% ## #
#,&# #$%#&#% "#
&#%,
&#
! 3 ""&# $
$ ""##$-$$, 0,#
&# # $/
5,
60"#, 76#$-,3"##$,
8,,&# 0$ /
6,#0##$,#2 $ "
5
6$ , 0 &# , #0 1$
6$ # "
69# "%&# "&##
# 0
6$! &#, , # # %
-2,2, %/
6#9#4,#&# $ 0
5"
6% #-#""/1 # # 0
# ,: -"*## 3 ,#/
6$ "",#,3&#,#&#
,#01 ,# #
http://163.16.28.248/bio/activelearner/

;"#, 76#$
6##, , 7&#"
6&# #$ 2 "%0,#,0#&#
,
6,3,###$,# , 0#&#%&#
6#$ ,# 0 "-( )/,
%!# 0 , 76#$-/
6< &#,# "< ",##$,#
http://www.ustboniface.mb.ca
,2#,# =>
?1=>
=@
,
5ABCDBC
&#,
EF
>0# ,$ ,### ###
#
.6
GH
.
GH
"#, ""2 ,#
0.IJ0-KL/
M0#&## ,#
+"K%#(##N)
6I.KJ7,-6
K
8A/
6K 0#0&#.0#,67,
"# 0
#.IOKP
QOLP
R
",#9##
+", ,

5#0 ", ,#%#0
5#0#, "
6>OKP.OK
>PSI
=8T
.OK
>P8TI
6=OKP.OK
>P7,-6I/
=8T
. T8I
! %
7, &# %,$ &# #$
MM.I
TOP
M0.I
=OPOP
M.-I
6=
HI
T/OP
K "OP..-OPOP/8U
U.-I
6=
HI
T/8I
=, " "<##$,#
M0OP
6OPVVU
M.-M
KW
OP/8U
6OP.U
M.M
KW
8T
6OPXXU
M.M
KW
# &#"M
KW
#U
# "< &#I
KA
.M
KW
8OPOP.OPHOP
6I
KA
##,-
6=
/
6" 0 &# "$ " &# &#" #, #$
6=8I
KA
# " &#
;#Y#N
,D 0%##$ 9#"KK,
6,!#,D,#-T,# ,, #Z,#
,&# "4 [Z,# , /
6%,D,#$!# $
6,,#, !,
,# # # !,%,%!%
,# Y#7 # !$$8$&#
A,## - "K%#/#$
## "
6,#,#2?@\
67 ,#, ,%&#
0##,$ ,# #
6: 0
6##-,&# /
6# #0&#
%$&#
+, #,,#4%$
$#7#7&#%$#
K # ""7 %$#
%$$ , #!0- , ",/
6,%$ 67-: "Y/
%$$ ,#, 78%$#
6%$,,, 7
$ 1 ",4%37-,, #/

#OP
#U
M
KW
,#
6%$,37##,! 37
,, , #
##%0
UM
KW
#
, ,##
http://blog.dearbornschools.org
5FC+KAAC5AKL+
#!'
7, # 2#1,'
4
4$#-# 2,3#$/
7, 667
#
K,, 7, %,!
K,37"#,# #, "Q=6,% 0##
1 "",%!
: ,3 , 2 2]6$7 2#$&#
6,,%,% HHH
0#,,%,%] "KA##D2%#
9#T%? %!
3 "&# $-HT[I^8 /
, 3,,I
# "#2#&# &#,#37#
�N$
6##&#
6,%
6%0 ,"
60# $

6T ! &#!U!M
6()-A2U
0/( ,)-2U
0$ /
637##$0 "
637# #0#, #:&#"##
: ,
7, # #
F # ,, #" #
F #I', # -02# /
I# ##0, . !
F !0 #,#,%,%
# " ,%,%
60$ # 2# "
6'# &#
63,6# -,%,% 2 2 #_/
# # ",# #6420 "%# #,# "
#7,,%,% $
# $90.0##FZ## ## "'
6#,9,%,% $
6#$# "7 0$,K
6, "7 $" 0,
67!FZ##1 0A
0$90.
6`>a ,%,% ##
6$ 1 A
6A,37 #
6$ 1 "K
6%,,###7%&#
64
1
/
5
100%