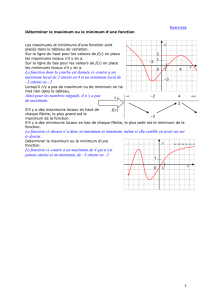
Les maximums profonds de silice biogénique

Les maximums profonds de silice biogénique : stratégie adaptative des diatomées face à
l’oligotrophisation?
Bernard QUÉGUINER, Sandrine CHIFFLET, Stéphanie JACQUET, Karine LEBLANC, Paul TRÉGUER
La sédimentation rapide des efflorescences de diatomées hors de la couche de surface, en
raison de la limitation en nutriments, est souvent considérée comme la règle générale dans les
écosystèmes océaniques. Plusieurs études réalisées au cours des deux dernières décennies ont aussi
montré l’existence de populations de diatomées en profondeur, formant des maximums profonds de
silice biogénique (DSMs pour Deep Silica Maximums) parfois associés aux maximums profonds de
chlorophylle (DCMs pour Deep Chlorophyll Maximums). Ces communautés caractérisent des eaux
limitées en surface par la disponibilité de nutriments (métaux–traces dans la zone HNLC de l’Océan
Austral, nutriments majeurs dans les zones oligotrophes). Le fonctionnement de ces écosystèmes
profonds, situés à la base de la couche océanique mélangée, doit faire appel à des mécanismes
particuliers couplant notamment le contrôle de la sédimentation, la résistance au broutage, les
particularités de la nutrition minérale des espèces dominantes, leur adaptation aux faibles
éclairements et les processus physiques contrôlant la transition entre les zones épi– et
mésopélagique.
Les observations réalisées dans l’Océan Austral (Quéguiner, 2001 ; Holm–Hansen & Hewes,
2004), en mer des Sargasses (Krause et al., 2009), dans le Pacifique Nord central et équatorial
(Blain et al., 1997; Brzezinski et al., 1998; Scharek et al., 1999, Dore et al., 2008) ainsi qu’en
Méditerranée (Crombet et al., 2011) montrent que les DSMs sont des structures biogéochimiques
répandues, constituées à la fois de silice biogénique détritique et de diatomées vivantes et présentes
au sein des nutriclines océaniques. Les assemblages sont dominés par des espèces fortement
silicifiées en fin de période estivale dans l’Océan Austral tandis que les études réalisées en
Méditerranée suggèrent une succession saisonnière marquée par une transition entre petites
diatomées peu silicifiées et grandes diatomées fortement silicifiées au cours de la saison.
Des études récentes réalisées en Arctique (Martin et al., 2010) suggèrent par ailleurs que les
DCMs sont également constitués par des populations de diatomées. Ces structures sont une
caractéristique dominante dans l’écosystème pélagique arctique mais elles restent fortement sous–
échantillonnées.
D’un point de vue conceptuel, les DSMs et DCMs observés dans les différentes régions
mentionnées sont expliqués par un développement des diatomées au sein d’une nutricline associée à
la pycnocline à une profondeur à laquelle la photoadaptation est possible pour les organismes
dominants. Leur persistance, malgré des taux de croissance vraisemblablement faibles, doit aussi
faire appel à des propriétés particulières de résistance au broutage par le zooplancton. Le devenir de
cette biomasse en fin de saison productive est particulièrement intéressant à étudier car il pourrait
s’agir d’une voie importante, bien qu’épisodique, de transfert de carbone organique vers le
compartiment mésopélagique. Le changement climatique doit s’accompagner d’un renforcement de
la stratification des eaux de surface, marqué par une diminution de l’épaisseur de la couche
mélangée océanique, ainsi qu’une diminution de la restauration hivernale des nutriments
(oligotrophisation). Les structures profondes pourraient alors constituer un refuge pour les
organismes phytoplanctoniques et la diminution d’épaisseur de la couche mélangée pourrait même
augmenter la productivité de ces systèmes par le biais d’une augmentation du rayonnement
disponible. Une attention particulière devrait donc être portée, en particulier dans le cadre du
chantier « Arctique », à l’étude du fonctionnement biologique et biogéochimique de ces maximums
profonds de biomasse en zone pélagique.
1
/
1
100%