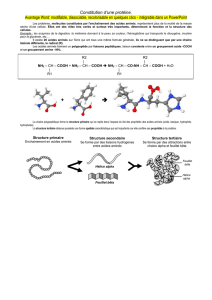
aminés polaire

Chapitre 6. Faisons le point sur la structure des protéines
6.1. Structures primaires, secondaires, tertiaires et quaternaires
6.2. La notion de domaine fonctionnel est essentielle
6.3. Les éléments de bases des structures secondaires, tertiaires et quaternaires
6.4. L ’hélice alpha et les autres hélices
6.5. Feuillets béta
6.6. Boucles et coudes
6.7. Les structures super-secondaires sont des motifs retrouvés dans de nombreuses protéines
6.8. Les éléments de bases des protéines : les acides aminés

6.3. Les éléments de base de la structure des protéines
6.3.1. La liaison peptidique est le ciment de base de toutes les structures protéiques (1)
Rappel : la structure commune de (presque) tous les acides aminés peut s’écrire :
R = chaîne latérale
COOH = fonction acide
NH2 = fonction amine
Le carbone central s’appelle Cα
-C -
R
NH2H
COOH
La liaison peptidique est une liaison covalente qui se forme par condensation du groupe
α-carboxyle (acide) d’un acide aminé avec le groupe α-aminé d’un autre acide aminé
et élimination d ’eau
liaison peptidique
(liaison amide particulière)
-C -C
R1
NH2
H
-C -COOH
R2
N
HO
H
H2O
+
+
-C -COOH
R1
NH2
H
-C -COOH
R2
N
H
H
H
α-carboxyle
α-amine

6.3.1. La liaison peptidique est le ciment de base de toutes les structures protéiques (2)
La liaison peptidique est une liaison amide particulière (W)
Elle est un « hybride de résonance » entre deux formes extrèmes
Première forme extrème
La liaison C O est une double liaison
La liaison C N est une simple liaison
L’azote de l’amide possède
une paire d’électrons non partagés
Forme hybride
Les électrons sont partagés
entre les atomes O, C et N
et sont distribués sur une
orbitale moléculaire π
qui recouvre les 3 atomes
Seconde forme extrème
La liaison C O est une simple liaison
La liaison C N est une double liaison
L’oxygène de l’acide possède
une paire d’électrons non partagés
En conséquence, la liaison peptidique possède trois propriétés fondamentales
Elle est plane Elle est rigide Elle est polaire

6.3.1. La liaison peptidique est le ciment de base de toutes les structures protéiques (3) (W)
La liaison peptidique présente des dimensions pratiquement fixes
carbone
azote
oxygène
hydrogène
Chaîne latérale
1,23 A
1,52 A
1,0 A
1,45 A
1,33 A
Cα
Cα
Il existe cependant 3 angles qui peuvent prendre des valeurs variables :
Cα
Cα
121,1°
123,2°
115,6°
119,5° 118,2°
121,9°
Ω
Ω
Ψ
Ψ
Φ
Φ

6.3.2. La liaison peptidique et la nature des résidus amino-acides impose des structures
spatiales très particulières aux chaînes polypeptidiques (W)
On définit Ω, angle de torsion autour de la liaison C-N. Cet angle ne peut prendre que 2 valeurs
Vers le
N-terminal
Vers le
C-terminal
Vers le
C-terminal
Vers le
N-terminal
Ω= 0° Ω= 180°
CIS TRANS
La liaison peptidique prend presque toujours une configuration trans, plus stable
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
1
/
28
100%