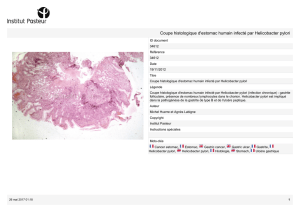
Mise en évidence de Helicobacter pylori à partir des biopsies

République Algérienne Démocratique et Populaire
Ministère de l’Enseignement Supérieur et de la Recherche Scientifique
Université d’Oran
Faculté des Sciences
Département de Biologie
Pour obtenir le diplôme de Docteur ES SCIENCES
en Biologie
Spécialité Microbiologie
Intitulé
Présenté par
Mme : Sous direction du :
MEDOUAKH Lynda
Pr BENSOLTANE Ahmed
.
Devant le jury :
Président AOUES Abdelkader
Professeur Université d’Oran
Directeur de thèse BENSOLTANE Ahmed Professeur Université d’Oran
Examinateur LAROUS Larbi Professeur Université de Sétif
Examinateur DJIBAOUI Rachid
Maitre de conférence -A- Université de Mostaganem
Examinateur CHOUGRANI Fadéla
Maitre de conférence -A- Université de Mostaganem
Examinateur BEKADA M. Ahmed
Maitre de conférence -A-
Université d’Oran
Année : 2010 - 2011
M
M
i
i
s
s
e
e
e
e
n
n
é
é
v
v
i
i
d
d
e
e
n
n
c
c
e
e
d
d
e
e
H
H
e
e
l
l
i
i
c
c
o
o
b
b
a
a
c
c
t
t
e
e
r
r
p
p
y
y
l
l
o
o
r
r
i
i
à
à
p
p
a
a
r
r
t
t
i
i
r
r
d
de
es
s
b
bi
io
op
ps
si
ie
es
s
g
ga
as
st
tr
ri
iq
qu
ue
es
s
e
et
t
s
so
on
n
a
an
nt
ta
ag
go
on
ni
is
sm
me
e
a
av
ve
ec
c
l
le
es
s
l
la
ac
ct
to
ob
ba
ac
ci
il
ll
le
es
s

Sommaire
Résumé
Liste des tableaux
Liste des figures
Liste des abréviations
Revue bibliographique
Introduction………………………………………………...........................................................................................1
I Généralité sur H. pylori
I-1 Historique ………………………….……………………………………………………….
………………………………………………4
I-2 Anatomie et Histologie…………………………………………………………………….…………………………………………..6
I-2-1 Anatomie……………………..…………………………………………………………..........……………………………………..6
I-2-2 Histologie………………………………………………………………………..………………………………………..……………….7
I-3 Physiologie de l’estomac……………………………………..………………………………………………………………………. 9
I-3-1-Fonction mécanique………………………………..…………………………………………………………………………………9
I-3-2-Fonction chimique…………………………………………………………………………………………..………………..........9
I-4 Mécanisme de régulation…………………………………………………………………………………………………………..10
I-5 Taxonomie de H. pylori …………………………………………………………..…………………………………….……………10
I-6 Ecologie………………………………………………………………………..………………………………………………….………...12
I-7 caractères morphologiques………………………………………………………………….........................................16
I-8 Caractères culturaux…......................................................................................................................17
I-9 caractères phénotypiques .........................................................................................................…...17
I-10 Caractères génétiques…………………………………………………………………………………………………………..…..19
I-11 Transmission…………………………………………………………………………......................................................21
I-12 Prévalence…………………………………………………………………………………………………………………….………….22
I-13 Facteurs de colonisation ………………………………………………………………………………………………….……….24
I-13-1 Mobilité …………………………………………………………………………………………………………..…………………….25
I-13-2 l’Urease …………………………………………………………………………………………………………………………..…….25
I-13-3 Adhérence ………………………………………………………………………………………………………………….…..…….25
I-14
M
ECANISME DE PERSISTANCE
………………………………………………………………………………………………………....26
I-15
R
EACTIONS INFLAMMATOIRES
…………………………………………………………………………………………………………..26
I-16
M
ÉTHODES DE DIAGNOSTIC
……………………………………………………………………………………………………….…...28
I-16-1
M
ETHODES INVASIVES
……………………………………………..………………………………………………………………….28
I-16-1-1
T
EST À L
’
URÉASE
……………………………………………………………………………………………………………………28
I-16-1-2
E
XAMEN CYTOLOGIQUE
…………………………………… ………………………………………………………………………31
I-16-1-3
E
XAMEN ANATOMOPATHOLOGIQUE
…………………………………………………………………………………………....31
I-16-1-4
CULTURE
…………………………………………………………………………………………………………………................32
I-16-1-5
PCR
EN TEMPS REEL
………………………………………………………………..………………………..……………….......32
I-16-2
M
ETHODES NON INVASIVES
………………………………..……………………………..………………….………………………33
I-16-2-1
T
ESTS RESPIRATOIRES
……………………………………….………………………………..……………..…………………….33
I-16-2-2
S
EROLOGIE
…………......................................................................................................................34
I-17
P
ATHOLOGIES ASSOCIEES A
H
ELICOBACTER PYLORI
………………………………………………..………….……………….…35
I-17-1
GASTRITES
…………………………………………………….…………………………………………………..………………………35
I-17-1-1
G
ASTRITE AIGUE
………………………………….…………………………..…………………………………..………………...37
I-17-1-2
G
ASTRITE CHRONIQUE
……………………………..…….…………………………………………………………..…………..
37
I-17-2
U
LCÈRE DUODÉNAL
…………………………………………………………………………………………………………………….
38
I-17-3
U
LCÈRE GASTRIQUE
………………………………………………………………....……………………………………………..…39
I-17-4
C
ANCER GASTRIQUE
………………………………………….……………………………..………………………………………….40
I-17-5
L
YMPHOME GASTRIQUE DE
MALT
………………………………………………………..………………..………………..….41
I-18
S
ENSIBILITE AUX ANTIBIOTIQUES
…………………………..……….………………………………..………………………………..43
I-19 Traitement………………………………………………………………………………………………………………………………..43
I-20 Traitement par les probiotiques ……………………………………………………………………………………………… 46
I-21 Vaccination………………………………………………………………………………………………………………………………..47
II- Généralité sur les bactéries lactiques………………………………………………………………………….…………......48
II-1 Historique……………………………………………………………………………………………………………………….………... 48
II-2 Caractères généraux ……………………………………………………………………….............................................48

II-3 Taxonomie ………………………………………………………………………………………………………………………………...50
II-4 Habitat ……………………………………………………………………………………....................................................52
II-5 Ecosystème gastro-intestinal……………………………………………………………….........................................52
II-6 Utilisation des bactéries lactiques dans le domaine agro-alimentaire …........................................55
II-7 Intérêts des bactéries lactiques en santé humaine ………………………………………...............................59
II-8 Souches probiotiques utilisées…………………………………………………………… ………………………………….. .59
II-9 Probiotiques et les infections gastro-intestinales……………………………………………………………............60
II-10 Interaction entre les bactéries lactiques…………………………………………………………………………….…...60
II-10-1 Inhibition des bactéries lactiques…………………………………………………………………………………….…….61
II-10-1-1 Acides organiques……………………………………………………………………….…………………………………....61
II-10-1-2 peroxyde d'hydrogène ……………………………………………………………………………………………………. 64
II-10-1-3 dioxyde de carbone ………………………………………………………………………………………………….........65
II-10-1-4 Diacétyle……………………………………………………………………………………………………………………………65
II-10-1-5 Acétaldéhyde………..…………………………………………………………………………………………………………..66
II-10-1-6 Reutérine……………………………………………………………….…………………………………………………………..66
II-10-1-7 Bactériocines …………………………………………………………………………………………………………………..67
II-11 Les lactobacilles………………………………………………………………………………………………………………………..68
II-11-1 Caractères généraux……………………………………………………………………………………………….…………….68
II-11-2 Habitat………………………………………………………………………………………………………………..…………………71
II-11-3 Effets des lactobacilles sur la santé…………………………………………….……………………………………71
Matériel et méthodes
Objectif du travail………………………………………….……………………………………………………………………………….74
I-Isolement et identification de H. pylori …………………………………………………………………..…………………….75
I-1 Echantillon et origine du matériel……………………………………….……………………………………………………..75
I-2 Prélèvement ……………………………………………………………………………………………………………………………...75
I-3 Conservation des biopsies …………………………………………………………………………………………….…………76
I-4 Examen anatomopathologique …………………………………………….…………………………………………………..76
I-5 Examens microbiologiques …………………………………………………………………………………………………..….. 76
I-5-1 Examen cytologique ………………………………………………………………………………………………….……….... 76
I-5-2 Test rapide à l’Urée……………………………………………………………………………..………………………………...77
I-5-3 Culture et isolement ……………………………………………………………………………..…………………...........7
8
I-5-3-1 Condition de culture ……………………………………………………………………………………..………………...78
I-5-3-2 Milieux de culture ……………………………………………………………………………………………………………..78
I-5-3-3 Isolement ………………………………………………………………………………….……………………………………...79
I-5-3-4 Identification ………………………………………………………………………………..…………………………………...79
I-5-3-4-1 Examen macroscopique…………………………………………………….……………………………………………….79
I-5-3-4-2 Examen microscopique …………………………………………………..…………………………………………………80
I-5-3-4-3 Etat frais………………………………………………………………………………………………………………………….…80
I-5-3-4-4 Tests biochimiques ………………………………………………………………………………………………………… 80
I-5-4 Antibiogramme ……………………………………………………………………………………………………………………….80
I-5-5 Conservation ..………………………………………………..………..…………………………………….……………………..81
I-5-6
A
PPLICATION DE LA TECHNIQUE
PCR
EN TEMPS REEL
………………………………………………………………………
83
II-
I
SOLEMENT ET CARACTERISATION DES LACTOBACILLES
…………………………………………………………………………......85
II-1
P
ROVENANCES DES ECHANTILLONS DU LAIT
…………………………………………………………………….…………………...85
II-2
M
ILIEUX DE CULTURE
……………………………………………………………………………….…………………………………….85
II-3
I
SOLEMENT ET IDENTIFICATION DES LACTOBACILLES
……………………………………………………………………………….85
II-3-1
I
SOLEMENT
……………………………………………………………………………………….……………………………………...85
II-3-2
I
DENTIFICATION DES LACTOBACILLES
……………………………………………………………..……………………………...86
II-3-2-1
C
ARACTERISATION MORPHOLOGIQUE
……………………………………………………..…………………………………....86
II-3-2-2
D
ETERMINATION DES ISOLATS
………………………………………………………................................................86
II-3-2-3
D
ETERMINATION DU GENRE
………………………………………………………………………………………………....…..87
II-3-2-3-1
T
EMPERATURE
………………………………………………………………
…………………………………………………..
87
II-3-2-3-2
T
YPE FERMENTAIRE
………………………………………………………………………………………………………………87
II-3-2-3-3
H
YDROLYSE DE L
’
ARGININE
……………………………………………………………………………………………………..87
II-3-2-3-4
U
TILISATION DU CITRATE
……………………………………………………………………………………………..…………88
II-3-2-3-5
H
YDROLYSE DE L
’
ESCULINE
………………………………………………………………….…………………………………88

II-3-2-4
D
ETERMINATION DE L
’
ESPECE
……………………………………………………………………..…………………………….88
II-3-2-4-1
I
DENTIFICATION PAR LE SYSTEME
API
50
CHL……………………………………………………………………………89
II-3-3
C
ONSERVATION DES SOUCHES
…………………………………………………………………….…………………………………90
III-
M
ISE EN EVIDENCE DES INHIBITIONS ENTRE LACTOBACILLE ET
H.
PYLORI
……………………….............................91
III-1
M
ETHODE DIRECTE
……………………………………………………………………………………………………………………….91
III-2
M
ETHODE INDIRECTE
……………………………………………………………………………………………………………………92
III-3
D
ETERMINATION DE LA NATURE DE L
’
AGENT INHIBITEUR
……………………………………………………………………….93
III-3-1
I
NHIBITION DUE AU PEROXYDE D
’
HYDROGENE
……………………………………………………………………………….93
III-3-2
I
NHIBITION DUE A L
’
ACIDE LACTIQUE
……………………………………………………………………………………………..93
III-3-3
I
NHIBITION DUE AUX BACTERIOPHAGES
………………………………………………………………………………………….94
III-3-4
R
ECHERCHES DE LA NATURE PROTEIQUE DE LA SUBSTANCE ANTIMICROBIENNE
…………………………………….94
III-3-5
E
FFET DE LA TEMPERATURE
…………………………………………………………...................................................95
III-4
M
ESURE DE L
’
ACIDITE TITRABLE
………………………………………………………………………………………………………..95
III-4-1
D
OSAGE DE L
’
ACIDITE
…………………………………………………………………………………………………………………95
III-5
M
ISE EN EVIDENCE DES INHIBITIONS EN MILIEU LIQUIDE
…………………………………………….………………………..96
III-6
E
FFET INHIBITEUR DES LACTOBACILLES SUR LES BACTERIES PATHOGENES
……………………….........................97
R
ESULTAT
I-M
ISE EN EVIDENCE DE
H.
PYLORI
………………………………….…………..…………………..……………………………………..98
I-1
E
XAMEN CYTOLOGIQUE
…………………………………………………………………………………………………………………….98
I-2
T
EST RAPIDE A L
’
UREE
………………………………………………………………….…………………………………………………..98
I
-3
E
XAMEN ANATOMOPATHOLOGIQUE
………………………………………………………..……………………………………….100
I
-4
I
SOLEMENT ET IDENTIFICATION
………………………………………………………..……………………………………………….100
I-4-1
A
SPECT MACROSCOPIQUE
……………………………………………………………..……………………………………………100
I-4-2
A
SPECT M
I
CROSCOPIQUE
……………………………………………………………………………………………..……………100
I-4-3
E
TAT FRAIS
……………………………………………………………………………………………………………………………….104
I-4-4
E
TUDE BIOCHIMIQUE
…………………………………………………………….……………………………………………………104
I-5
A
NTIBIOGRAMME
…………………………………………………………………………………………………………………………105
I-6
M
ISE EN EVIDENCE DE
H.
PYLORI PAR LA
PCR
EN TEMPS REEL
………………………………………………………….…….
109
II
I
SOLEMENT ET IDENTIFICATION DES LACTOBACILLES
…………………………………………………………………………………115
II-1
E
XAMEN MORPHOLOGIQUE
…………………………………………………………………………………………………………...115
II-2
C
ARACTERES PHYSIOLOGIQUES ET BIOCHIMIQUES
………………………………………………………………………………..115
II-3
R
EPARTITION DES ESPECES DE LACTOBACILLE
………………………………………………………………………..……………127
III
P
RODUCTION DES SUBSTANCES ANTIMICROBIENNES
………………………………………………………………………….....129
III-1 Mise en évidence des inhibitions entre les lactobacilles et H. pylori …………………………………….…129
III- 2 Acidification……………………………………………….…………………………………………………………………………..139
III-3 Détermination de l’agent inhibiteur ………………………………………..……………………………………………..143
III-3-1 Effet de la production d’acide lactique ………………………………………………………………………………..144
III-3-2 Effet du peroxyde d’hydrogène ……………………………………………………………………..……………………146
III-3-3 productions de phage lysogène: ………………………………………………………………………………………….146
III-3-4 Effet de la bactériocine…………………..……………………………………………………………………….…….…... 147
III-4 Effet de surnageant de lactobacille sur H. pylori en milieu liquide ………….…………………………....150
III-5 Inhibitions des bactéries pathogènes par les lactobacilles…………………………………………………..…..152
Discussion ……………….………………………………………………………………………………………….…………………………156
Conclusion et perspectives….………………………………............................................................................179
Références bibliographiques ….……………………………………………………………………………..………………………182
Annexe……………………………………………………………………………………………………………................................215
Activités scientifiques…………………………………………………………………………………………………………………….219

Résumé
Le but de ce présent travail est de détecter in vitro le pouvoir inhibiteur des souches de
Lactobacilles isolés du lait de chèvre sur la bactérie pathogène de H. pylori isolée des biopsies
gastriques et de déterminer l’agent inhibiteur.
En premier temps, la recherche de H. pylori a été réalisé à partir des biopsies gastriques des
patients atteints de maladies gastroduodénales (gastrite chronique, ulcére gastroduodénale), et
ayant subi une endoscopie digestive haute.
La présence de cette bactérie a été mise en évidence par des tests invasifs : test rapide à
l’uréase, l’examen cytologique, l’examen histologique, la culture et l’amplification génique
PCR en temps réel.
L’identification des souches de H. pylori isolées a été basée sur les caractères
morphologiques (macroscopique et microscopique) et biochimiques (test à l’uréase, test à
l’oxydase et la catalase).Nous avons pu isoler trois souches de H. pylori alors que les autres
tests invasifs utilisés montrent parfois la présence de H. pylori dans les échantillons présentant
une négativité de la culture (résultat faussement négatif). Ceci est probablement du à l’erreur
de l’échantillonnage ou la sensibilité de la culture. .
Par ailleurs, l’étude de la sensibilité des souches de H. pylori aux antibiotiques habituellement
utilisés dans le choix thérapeutique a montré l’excellente activité de la plupart des
antibiotiques testés vis-à-vis de H. pylori. Aucune résistance à la clarithromycine n’a été
observée chez toutes les souches de H. pylori, cette résistance constitue le facteur majeur
d’échec du traitement d’éradication de H. pylori.
De plus, La technique moléculaire de la PCR en temps réel a permis de détecter à la fois H.
pylori et d’éventuelle mutation de résistance à la clarithromycine à partir des biopsies
gastriques ou de la colonie. En effet, les souches isolées de H. pylori (HP1, HP2, HP3),
traitées par cette technique ont révélé d’une part leur conformité à l’Helicobacter pylori
(ADN positif). Elles sont identifiées comme des souches dites sauvages (Wite type), et d’autre
part, montre que ces souches ne présentent aucune mutation de résistance à la clarithromycine
En second temps, 42 souches de lactobacilles ont été isolés à partir du lait cru de chèvre, et
identifiés en se basant sur des critères morpholologiques,biochimiques et physiologiques. Les
souches lactiques sont rattachées aux 3 groupes (I, II, III). Par ailleurs, cette identification a
révélé la présence d’une diversité en espéces dans le genre de lactobacilles répartie
essentiellement entre Lb. delbrueckii (sp. bulgaricus et sp. lactis ); Lb. helveticus; Lb.
salivarius ; Lb. plantarum ; Lb. casei (sp. casei et sp. rahmnosus) ; Lb. rhamnosus ; Lb.
paracasei ; Lb. pentosus ; Lb. brevis et Lb. fermentum.
D’autre part, nous avons testé l’activité antimicrobienne in vitro des lactobacilles isolés ainsi
que la souche de référence vis-à-vis des souches isolés de H. pylori (HP1, HP2, HP3) et les
souches de référence (HP4) et (HP5) par la méthode directe de diffusion (Geis et al., 1983).
Les souches de Lactobacillus sp. ont présenté dans l’ensemble une activité antagoniste vis-à-
vis de H. pylori, démontrée par l'apparition de zone d’inhibition avec des diamètres variés
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
208
209
210
211
212
213
214
215
216
217
218
219
220
221
222
223
224
225
226
227
228
229
230
231
232
233
234
1
/
234
100%