the document - La bibliothèque numérique de la

Par Elias Narciso MATOS
African Virtual university
Université Virtuelle Africaine
Universidade Virtual Africana
Macromolécules
dans les systèmes
biologiques

Université Virtuelle Africaine 1
Note
Ce document est publié sous une licence Creative Commons.
http://en.wikipedia.org/wiki/Creative_Commons
Attribution
http://creativecommons.org/licenses/by/2.5/
License (abréviation « cc-by »), Version 2.5.

Université Virtuelle Africaine 2
I. Macromolécules dans les systèmes biologiques ___________________3
II. Cours et savoir-faire pré-requis _______________________________3
III. Temps ___________________________________________________3
IV. Matériels didactiques _______________________________________3
V. Justification_______________________________________________4
VI. Contenu__________________________________________________4
6.1 Résumé _______________________________________________4
6.2 Représentation graphique _________________________________ 4
VII. Objectifs généraux _________________________________________5
VIII. Objectifs spécifiques d’apprentissage ___________________________6
IX. Activités d’enseignement et d’apprentissage ______________________8
X. Activités d’apprentissage ___________________________________ 14
XI. Liste des concepts clés (glossaire) ____________________________90
XII. Liste des lectures obligatoires ________________________________93
XIII. Listes des liens utiles ______________________________________95
XIV. Synthèse du module _______________________________________ 99
XV. Évaluation sommative _____________________________________100
XVI. Références bibliographiques ________________________________106
XVII. Auteur principal du module ________________________________107

Université Virtuelle Africaine 3
Par Dr. Elias Narciso Matos.
L’étudiant doit connaître le contenu fondamental couvert dans le curriculum du
programme de chimie et, en particulier les sujets concernant principalement les
modules 5, 6, 7.
Notion d’isomérie et de stœchiométrie
Structure et réactivité des aldéhydes et des cétones
Structure et réactivité des acides aminés
Concept des aromatiques et des hétérocycles
120 heures
Unité Heures
Glucides 40
Protéines 40
Lipides 30
Activité pratique 10
Total 120
Le module 8 (macromolécules dans les systèmes biologiques) est basé sur le maté-
riel d’enseignement suivant : ouvrages scolaires de chimie, logiciel éducationnel de
chimie, expériences virtuelles réalisées par ordinateur, expériences simples, manuels
scolaires et manuels.

Université Virtuelle Africaine 4
Ce module introduit l’étude de quelques composés représentatifs des macromolé-
cules naturelles qui sont vitales pour l’équilibre, le développement, l’entretien et les
suppléments des organismes vivants.
Le module consolide et élargie les connaissances acquises à partir des modules précé-
dents, principalement sur les aldéhydes, les cétones et les acides aminés, en les reliant
aux fonctions vitales de l’organisme. L’étude du contenu de ce module apportera
une connaissance sur les propriétés et les activités de certaines des macromolécules
importantes pour la survie des organismes.
6.1. Résumé
Le présent module 8 (macromolécules dans les systèmes biologiques) comporte 4
unités qui sont : les glucides, les protéines, les lipides et nalement des activités
pratiques.
Chaque unité établie et analyse les connaissances fondamentales sur la structure,
la nomenclature et la synthèse, ainsi que les propriétés chimiques et physiques des
macromolécules ayant un intérêt biologique.
Pour l’aspect pratique, le module offre une formation sur comment manier les
équipements de base du laboratoire et/ou des exercices simulés par ordinateur sur
les techniques de synthèse, de séparation, de purication et de détermination des
constantes physiques d’un composé.
Le module dure au total 120 heures.
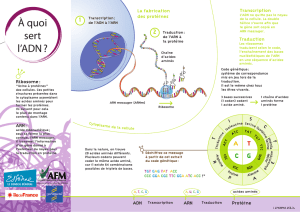
6.2 Représentation graphique
M acr omolécules
G lucides
Pr otéines
Lipid es
Macromolécules
Glucides
Protéines
Lipides
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
1
/
151
100%