Interaction cellule-matériau: une modulation - iPubli

NOUVELLES
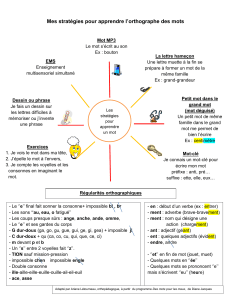
Interaction cellule-matériau:
une modulation biochimique ou biophysique 7
•
•
N)

.
:
1
/
2
100%