Enzymologie - WordPress.com

Enzymologie Générale
Structure et Propriétés des
enzymes

I. Généralités
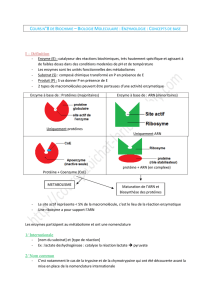
I.1. Définition
Les enzymes sont des biocatalyseurs protéiques
spécifiques qui accélèrent la vitesse des réactions
chimiques se déroulant dans la cellule.
Le pouvoir enzymatique est intimement lié à la
structure tridimensionnelle de la protéine
enzymatique, il est altéré quand cette dernière est
modifiée.

I.2. Propriétés générales des enzymes
Les enzymes ne créent pas la réaction, elles la
favorisent et sont complètement régénérées à la
fin de celle là.
La réaction est en général une rupture de
liaison covalente
Une formation de liaison de covalente
Une association des deux.

Les enzymes abaissent l’énergie d’activation et
augmentent la concentration des substrats à
l’état activé.
Les enzymes n’interviennent pas dans
l’équilibre des réactions chimiques réversibles.

Les enzymes agissent à de très faibles
concentrations, d’où la notion d’efficacité
déterminée par l’activité moléculaire spécifique
(AMS). L’activité moléculaire spécifique étant
définie comme le nombre de molécules de
substrats transformées par unité de temps et par
molécule d’enzyme, lorsque cette dernière est le
facteur limitant de la réaction : 103 AMS < 106.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
1
/
59
100%