Les héparines dans le traitement des cancéreux

Forum Med Suisse 2011 ;11(10):169–171 169
curriculum
Les héparines dans le traitement des cancéreux
Heinz Läublia,Lubor Borsigb
aKlinik Innere Medizin, Universitätsspital Basel
bPhysiologisches Institut und ZürichCenter für Integrative Humanphysiologie (ZIHP), Universität Zürich
Introduction
L’ incidence accrue des accidents thrombo-emboliques
chez les cancéreux aété décrite pour la première fois en
1865 par Armand Trousseau [1]. Ce phénomène est im-
putable àdes facteurs activateurs provenant de la tu-
meur elle-même ou de son microenvironnement (tumor
microenvironment) activé [2]. En plus de cela, plusieurs
modalités thérapeutiques anticancéreuses sont throm-
bogènes [3, 4]. Les 3mécanismes thrombogènes pos-
tulés par Virchow peuvent être déclenchés par la tu-
meur elle-même: perturbations hémodynamiques,
activation de l’endothélium et du système hémostatique.
La compression directe des vaisseaux peut provoquer
leur occlusion et du même fait des thrombo-embolies. Au
cours de la progression de la tumeur,des cytokines pro-
inflammatoires parviennent dans le sang et peuvent ac-
tiver l’endothélium local et systémique. La tumeur peut
en outre activer les thrombocytes et la cascade de la coa-
gulation. Plusieurs mécanismes moléculaires ont pu être
identifiés ces dernières années. Le tissue factor (TF,fac-
teur III), qui intervient dans l’activation de la cascade de
la coagulation et des thrombocytes, est exprimé directe-
ment sur les cellules tumorales et dans le microenviron-
nement de la tumeur.Avec la progression de la patholo-
gie tumorale, il se produit une libération de microparticules
de TF thrombogènes dans la circulation [5]. Le TF et l’in-
hibiteur de la plasmine PAI-1 sont induits par l’hypoxie
en suivant la voie de l’hypoxia inducible factor (HIF) [6].
Les mucines sont de grosses molécules dotées d’un
noyau protéique auxquelles sont liées par covalence de
longues chaînes polysaccharidiques.Ellesforment
normalement une couche protectrice des épithéliums et
sont sécrétées dans le sang par les tumeurs épithéliales.
Ces mucines intravasculaires peuvent provoquer des
thrombo-embolies directement ou par activation leuco-
cytaire [2].
L’ héparine est un glyco-aminoglycane naturel, donc un
polysaccharide, présent dans les mastocytes. L’ héparine
non fractionnée, isolée du tractus gastro-intestinal de
porc, est un mélange de plusieurs longues chaînes sac-
charidiques (200–300 monosaccharides alignés) [7]. Les
héparines de bas poids moléculaire sont extraites de
l’héparine naturelle et forment des chaînes polysaccha-
ridiques raccourcies par méthodes chimiques et phy-
siques. Pour l’activité sur l’antithrombine III, qui assure
l’effet anticoagulant des héparines, seul un petit poly-
mère de 5 sucres est nécessaire [8]. Cette séquence sac-
charidique minimale est obtenue par synthèse et est uti-
lisée en clinique sous le nom de fondaparinux (Arixtra®).
Héparines et survie
L’ étude CLOT publiée en 2003 a montré que les cancé-
reux traités par daltéparine (Fragmin®) avaient signifi-
cativement moins de récidives thrombo-emboliques
queceux traités par un antagoniste de la vitamine K[9].
6 mois après le début du traitement, 27 patients sur les
336 du groupe daltéparine ont eu un nouvel accident
thrombo-embolique, contre 53 sur les 336 du groupe
antagoniste de la vitamine K (HR 0,48). L’ incidence des
hémorragies a été la même dans les deux groupes. Les
analyses de survie de cette même étude ont en outre ré-
vélé une survie plus longue des patients traités par dal-
téparine et n’ayant aucune métastase clinique lors de
cette intervention (survie globale à 12 mois 80% contre
64% dans le groupe antagoniste de la vitamine K) [10].
Une autre étude de 84 patients ayant un carcinome
bronchique à petites cellules a comparé un traitement
par héparines de bas poids moléculaire à un placebo et
a montré une amélioration significative de la survie
sans progression de 6 à 10 mois [11]. Un effet a été
constaté aussi bien au stade limited disease qu’au
stade extensive disease.
Une étude effectuée dans les années 1990 déjà chez
277 patients ayant un carcinome bronchique à petites
Vous trouverez les questions à choix multiple concernant cet article
à la page 168 ou sur Internet sous www.smf-cme.ch.
Heinz Läubli
Les auteurs
certifient
qu’aucun conflit
d’intérêt n’est lié
à cet article.
Quintessence
PLes accidents thrombo-emboliques sont plus fréquents chez les cancé-
reux. La thrombophilie peut être secondaire àlatumeur elle-même ou à
son traitement.
PDans l’étude CLOT, le traitement des cancéreux thrombo-emboliques
par héparine de bas poids moléculaire est supérieur à celui par antago-
nistes de la vitamine K.
PEn plus de cette supériorité dans le traitement des accidents thrombo-
emboliques, les études cliniques ont révélé un effet antinéoplasique des
héparines.
PL’ effet antinéoplasique des héparines repose sur différents mécanis-
mes biologiques. Les sélectines (molécules d’adhésion cellulaire vascu-
laire) notamment sont inhibées, qui ont une importante fonction lors de
la métastatisation. L’ effet antimétastatique des héparines est indépen-
dant de leur activité anticoagulante et ne semble important qu’à la phase
précoce de la métastatisation.
PDe nouvelles études cliniques avec des héparines à des stades tumo-
raux précoces sont en cours. De plus, de nouvelles héparines modifiées à
effet spécifique sur les sélectines sont en phase préclinique.

Forum Med Suisse 2011 ;11(10):169–171 170
curriculum
cellules a montré dans le groupe intervention, qui a
reçu en plus du traitement standard une héparine non
fractionnée sous-cutanée pendant 5 semaines, une
amélioration de la survie moyenne (317 jours contre
261 pour le placebo) [12]. Dans l’étude FAMOUS chez
385 patients ayant un cancer à un stade avancé, Kak-
kar et ses collègues ont trouvé un avantage de survie si-
gnificatif à 17 mois dans le groupe ayant reçu pendant
1 année 5000 U.I. de daltéparine sous-cutanées au lieu
du placebo [13]. Un groupe de 148 patients à un stade
avancé de leur cancer a eu un avantage de survie signi-
ficatif contre placebo sous traitement pendant 1 an par
nadroparine (Fraxiparine®) à dose prophylactique [14].
Comparativement à cela, les 154 patients ayant reçu un
placebo sont décédés 1,33 fois plus souvent à 1 an.
Dans une petite étude, Sideras et ses collègues n’ont par
contre constaté aucun effet significatif de la daltéparine
sur la survie de cancéreux [15]. Un aperçu systéma-
tique des résultats des études est donné dans le travail
de Kuderer et collègues [4].
Effet antinéoplasique des héparines
L’ effet des héparines sur la survie des cancéreux s’ex-
plique en partie par la prophylaxie des thrombo-embo-
lies, avec leur morbidité et leur mortalité. Les patients
sous traitements thrombogènes tels que thalidomide
ou lénalidomide devraient donc recevoir une héparine
quand possible et même en ambulatoire [16]. Certains
résultats d’études cliniques et expérimentales montrent
cependantqu’il yauneffet antinéoplasique direct. L’ ex-
périmentation animale a montré à plusieurs reprises
que l’héparine non fractionnée et les héparines de bas
poids moléculaire ont une influence sur la progression
tumorale et la métastatisation [17, 18]. En plus de l’effet
sur la coagulation par la liaison de la séquence pentasac-
charidique àl’antithrombine III, l’héparine non fraction-
née, et de manière différente les héparines de bas poids
moléculaire, ont une influence sur de nombreux méca-
nismes biologiques.
L’ héparine non fractionnée et quelques héparines de
bas poids moléculaire se lient à des molécules d’adhé-
sion cellulaire telles qu’intégrines et sélectines et inhi-
bent de ce faitleurs interactions [7, 19, 20]. Des chimio-
kines et facteurs de croissance sont également liés par
les héparines [7, 21]. Fait intéressant, la liaison aux
chimiokines telles que le platelet factor 4 (PF4) est
fonction de la longueur de la chaîne polysaccharidique.
PF4 plus héparine sont l’antigène dans la thrombo-
pénie immunitaire induite par l’héparine (HIT). Ce qui
signifie que contrairement aux longues héparines le
pentasaccharide fondaparinux ne provoque pas d’HIT
et peut donc être utilisé chez les patients ayant des
anticorps HIT. Les enzymes liant les héparanes tels que
l’héparanase sont inhibés par les héparines [22]. La sé-
crétion de tissue factor pathway inhibitors (TFPI) peut
en outre être influencée par les héparines [23]. Bien
que théoriquement tous ces mécanismes puissent jouer
un rôle, il a été démontré dans le modèle de souris que
c’est surtout l’inhibition des interactions des sélectines
pendant la métastatisation hématogène qui conditionne
l’effet antinéoplasique des héparines in vivo.
Sélectines et niche métastatique
Les modèles murins, mais surtout les modèles expéri-
mentaux de métastases par injection intraveineuse de tu-
meurs, ont pu montrer que les héparines ont un effet sur-
tout antimétastatique [24]. Pendant la métastatisation
hématogène, les cellules tumorales infiltrent les vaisseaux
sanguins directement ou par les canaux lymphatiques.
Dans le vaisseau, il se produit une interaction avec les
éléments sanguins cellulaires et acellulaires, surtout les
thrombocytes. Ce qui déclenche la formation d’agrégats
de cellules tumorales-thrombocytes qui peuvent libérer
des emboles dans la microvascularisation des organes. A
partir de ces emboles tumoraux, il se produit un condi-
tionnement du microenvironnement avec création d’un
voisinage permissif pour les cellules tumorales, ladite
niche métastatique (metastatic niche) dans laquelle les cel-
lules tumorales peuvent survivre, s’extravaser,infiltrer le
tissu, proliférer et former une nouvelle métastase [25, 26].
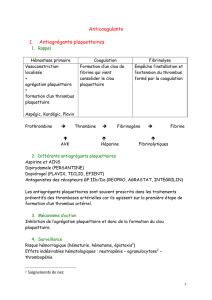
Figure 1
Cascade métastatique et mécanismes influencés par les héparines.
A: L’ effet des héparines sur les cytokines permet d’inhiber l’angiogenèse et consécutive-
ment aussi l’intravasation de cellules tumorales. Cette intravasation peut être inhibée
par l’effet des héparines sur l’héparanase. L’ inhibition de la sélectine P prévient en
outre la formation d’agrégats cellules tumorales-thrombocytes (emboles tumoraux).
B: Au niveau des petits vaisseaux des organes dans lesquels de nouvelles métastases
apparaissent, les héparines peuvent réduire l’activation du microenvironnement et
empêcher la création d’une niche métastatique par inhibition des sélectines P et L,
de l’activation des thrombocytes et de la cascade de la coagulation. Le recrutement
des leucocytes peut être partiellement bloqué par le fait que les héparines empêchent
les liaisons dépendant des intégrines et sélectines.
C: Après activation du microenvironnement et création d’une niche métastatique,
les cellules tumorales peuvent proliférer dans de nouveaux organes. Les héparines
peuvent ici aussi influencer la transmission de signaux par les cytokines.

Forum Med Suisse 2011 ;11(10):169–171 171
curriculum
Les sélectines sont des molécules d’adhésion cellulaire
vasculaire qui donnent des interactions hétérotypiques
avec les ligands saccharidiques par leur domaine lec-
tine. La sélectine P est stockée dans les granules des
thrombocytes et les corps de Weibel-Pallade des cellules
endothéliales, et après activation présentée à la surface
des thrombocytes ou des cellules endothéliales. La sé-
lectine E est synthétisée de novo après activation par
les cellules endothéliales. La sélectine L est exprimée
constitutionnellement sur tous les leucocytes myéloïdes
et la plupart des lymphocytes [27]. Les sélectines pro-
voquent le freinage initial des leucocytes pendant l’ex-
travasation lors des réactions inflammatoires et pen-
dant le trafficking normal, par ex. dans les organes
lymphatiques secondaires [28]. La sélectine Paenoutre
une influence directe sur l’adhésivité plaquettaire [29].
Les cellules des tumeurs épithéliales produisent des
mucines polysaccharidiques à leur surface et les libè-
rent dans le sang périphérique (voir Introduction). Les
mucinesàlasurface des cellules tumorales déclenchent
l’interaction avec la sélectine P sur les thrombocytes.
Cette interaction est à l’origine de l’embole tumoral
[24, 30]. Les sélectines P et L sont en outre respon-
sables de l’activation du microenvironnement, surtout
des cellules endothéliales, et permettent ainsi la créa-
tion d’une niche métastatique [31]. La sélectine L est en
outre impliquée dans le recrutement des leucocytes
myéloïdes, qui ont une fonction importante dans la
création de la niche métastatique et donc de nouvelles
métastases [30, 32, 33].
L’ inhibition des sélectines par héparines
empêche la métastatisation
Les sélectines contrôlent d’importantes étapes dans la
métastatisation hématogène des cellules tumorales [34].
L’ ensemencement expérimental des poumons par injec-
tion intraveineuse de cellules de cancer colique est net-
tement moins important chez les souris déficientes en
sélectine PouL[30]. Fait intéressant, il yaune synergie
d’effets, àsavoir que chez les souris doublement défi-
cientes en sélectine PetLles métastases sont encore
moins nombreuses que chez les monodéficientes. Ces ef-
fets ont pu être reproduits avec l’héparine non fraction-
née [24, 32]. Un traitement par héparine peu avant l’in-
jection intraveineuse de cellules tumorales adiminué la
métastatisation par un mécanisme dépendant de la sé-
lectine P[24]. Aucun autre effet n’a pu être observé chez
les souris déficientes en sélectine P. Mais si l’héparine a
été injectée 6–12 heures après l’injection de cellules tu-
morales àdes souris déficientes en sélectine P, une dimi-
nution plus marquée de la formation de métastases apu
être constatée [32]. Cet effet àretardement de l’héparine
semble dépendre de la sélectine Lcar il ne se produit pas
chez les souris qui en sont déficientes [32]. Chez les sou-
ris doublement déficientes en sélectine PetLetdans le
modèle expérimental de métastases utilisé, l’héparine
non fractionnée pendant les premières heures suivant
l’inoculation de cellules tumorales n’a montré aucune
diminution de la métastatisation [30]. Ces résultats indi-
quent que la propriété antimétastatique de l’héparine
non fractionnée est principalement due àl’inhibition des
sélectines et n’ad’importance qu’au cours des premières
heures aprèsinjection de cellules tumorales. Les hépa-
rines de bas poids moléculaire peuvent plus ou moins
bien lier et inhiber les sélectines et donc plus ou moins
bien aussi la métastatisation [35, 36]. Les héparines à
courte chaîne notamment, comme la nadroparine, ont
un effet moins marqué. Il faut savoir àcepropos que le
pentasaccharide fondaparinux n’a aucun effet sur cette
métastatisation expérimentale [35]. Ce qui permet de
conclure que l’effet antimétastatique des héparines n’est
pas dû àleur activité anticoagulante. Des expériences
avec héparines modifiées sans activité sur la coagulation
mais avec puissant effet inhibiteur des sélectines ont
déjà pu être réalisées avec succès dans un modèle mé-
tastatique expérimental [37].
Perspectives
Chez les cancéreux, les héparines préviennent une réci-
dive de thrombo-embolie plus efficacement que les an-
tagonistes de la vitamine K, du fait de leur activité bio-
logique plus large. Les cancéreux thrombo-emboliques
doivent donc si possible toujours être traités par hépa-
rines. En plus de la diminution des thrombo-embolies,
les héparines ont aussi un effet antinéoplasique direct,
essentiellement dû àl’inhibition des sélectines pendant
la métastatisation. Les études cliniques s’intéressant à
l’effet des héparines devraient donc se concentrer sur-
tout sur des stades tumoraux précoces, avant toute mé-
tastatisation. Cette population de patients aleplus pro-
fité de l’intervention par daltéparine dans l’étude CLOT
aussi [10]. Plusieurs études en cours examinent l’effet
des héparines de bas poids moléculaire chez des pa-
tients àunstade tumoral localisé. Des héparines modi-
fiées ayant une activité spécifique sur les sélectines par
ex. sont en outre testées dans des modèles précliniques.
Ces substances viendront compléter les traitements
antitumoraux aussi bien classiques que ciblés.
Correspondance:
Heinz Läubli
Klinik Innere Medizin
Universitätsspital Basel
CH-4031 Basel
Références recommandées
–Lee AY, Levine MN, Baker RI, Bowden C, Kakkar AK, Prins M, et al.
Low-molecular-weight heparin versus a coumarin for the prevention
of recurrentvenous thromboembolism in patients with cancer.NEngl
J Med. 2003;349:146–53.
–Borsig L, Wong R, Feramisco J, Nadeau DR, Varki NM, Varki A.
Heparin and cancer revisited: mechanistic connections involving
platelets, P-selectin, carcinoma mucins, and tumor metastasis. Proc
Natl Acad Sci. U S A 2001;98:3352–7.
–Joyce JA, Pollard JW. Microenvironmental regulation of metastasis.
Nat Rev Cancer. 2009;9:239–52.
–Psaila B, Lyden D. The metastatic niche: adapting the foreign soil. Nat
Rev Cancer. 2009;9:285–93.
–Läubli H, Borsig L. Selectins promote tumor metastasis. Semin Can-
cer Biol. 2010;20:169–77.
Vous trouverez la liste complète et numérotée des références dans la
version en ligne de cet article sous www.medicalforum.ch.

Heparine in der Behandlung von Tumorpatienten /
Les héparines dans le traitement des cancéreux
Literatur (Online-Version) / Références (online version)
1 Trousseau A. Phlegmasia alba dolens. Lectures on clinical medicine, delivered at the Hotel-Dieu,
Paris 1865;5:281–332.
2 Varki A. Trousseau’s syndrome: multiple definitions and multiple mechanisms. Blood.
2007;110:1723–9.
3 Lip GY, Chin BS, Blann AD. Cancer and the prothrombotic state. Lancet Oncol. 2002;3:27–34.
4 Kuderer NM, Ortel TL, Francis CW. Impact of venous thromboembolism and anticoagulation on
cancer and cancer survival. J Clin Oncol. 2009;27:4902–11.
5 Kasthuri RS, Taubman MB, Mackman N. Role of tissue factor in cancer. J Clin Oncol.
2009;27:4834–8.
6 Denko NC, Giaccia AJ. Tumor hypoxia, the physiological link between Trousseau’s syndrome
(carcinoma-induced coagulopathy) and metastasis. Cancer Res. 2001;61:795–8.
7 Lever R, Page CP. Novel drug development opportunities for heparin. Nat Rev Drug Discov.
2002;1:140–8.
8 Turpie AG, Gallus AS, Hoek JA. A synthetic pentasaccharide for the prevention of deep-vein
thrombosis after total hip replacement. N Engl J Med. 2001;344:619–25.
9 Lee AY, Levine MN, Baker RI, Bowden C, Kakkar AK, Prins M, et al. Low-molecular-weight
heparin versus a coumarin for the prevention of recurrent venous thromboembolism in patients
with cancer. N Engl J Med. 2003;349:146–53.
10 Lee AY, Rickles FR, Julian JA, Gent M, Baker RI, Bowden C, et al. Randomized comparison of
low molecular weight heparin and coumarin derivatives on the survival of patients with cancer
and venous thromboembolism. J Clin Oncol. 2005;23:2123–9.
11 Altinbas M, Coskun HS, Er O, Ozkan M, Eser B, Unal A, et al. A randomized clinical trial of
combination chemotherapy with and without low-molecular-weight heparin in small cell lung
cancer. J Thromb Haemost. 2004;2:1266–71.
12 Lebeau B, Chastang C, Brechot JM, Capron F, Dautzenberg B, Delaisements C, et al.
Subcutaneous heparin treatment increases survival in small cell lung cancer. “Petites Cellules”
Group. Cancer. 1994;74:38–45.
13 Kakkar AK, Levine MN, Kadziola Z, Lemoine NR, Low V, Patel HK, et al. Low molecular
weight heparin, therapy with dalteparin, and survival in advanced cancer: the fragmin advanced
malignancy outcome study (FAMOUS). J Clin Oncol. 2004;22:1944–8.
14 Klerk CP, Smorenburg SM, Otten HM, Lensing AW, Prins MH, Piovella F, et al. The effect of
low molecular weight heparin on survival in patients with advanced malignancy. J Clin Oncol.
2005;23:2130–5.
15 Sideras K, Schaefer PL, Okuno SH, Sloan JA, Kutteh L, Fitch TR, et al. Low-molecular-weight
heparin in patients with advanced cancer: a phase 3 clinical trial. Mayo Clin Proc. 2006;81:758–
67.
16 Brose KM, Lee AY. Cancer-associated thrombosis: prevention and treatment. Curr Oncol.
2008;15:S58–67.
17 Borsig L. Heparin as an inhibitor of cancer progression. Prog Mol Biol Transl Sci. 93:335–49.
18 Smorenburg SM, Van Noorden CJ. The complex effects of heparins on cancer progression and
metastasis in experimental studies. Pharmacol Rev. 2001;53:93–105.
19 Peter K, Schwarz M, Conradt C, Nordt T, Moser M, Kubler W, Bode C. Heparin inhibits ligand
binding to the leukocyte integrin Mac-1 (CD11b/CD18). Circulation. 1999;100:1533–9.
1

20 Koenig A, Norgard-Sumnicht K, Linhardt R, Varki A. Differential interactions of heparin and
heparan sulfate glycosaminoglycans with the selectins. Implications for the use of unfractionated
and low molecular weight heparins as therapeutic agents. J Clin Invest. 1998;101:877–89.
21 Lundin L, Larsson H, Kreuger J, Kanda S, Lindahl U, Salmivirta M, Claesson-Welsh, L.
Selectively desulfated heparin inhibits fibroblast growth factor-induced mitogenicity and
angiogenesis. J Biol Chem. 2000;275:24653–60.
22 Vlodavsky I, Ilan N, Nadir Y, Brenner B, Katz BZ, Naggi A, et al. Heparanase, heparin and the
coagulation system in cancer progression. Thromb Res. 120(Suppl 2):S112–20.
23 Mousa SA, Linhardt R, Francis JL, Amirkhosravi A. Anti-metastatic effect of a non-anticoagulant
low-molecular-weight heparin versus the standard low-molecular-weight heparin, enoxaparin.
Thromb Haemost. 2006;96:816–21.
24 Borsig L, Wong R, Feramisco J, Nadeau DR, Varki NM, Varki A. Heparin and cancer revisited:
mechanistic connections involving platelets, P-selectin, carcinoma mucins, and tumor metastasis.
Proc Natl Acad Sci. U S A 2001;98:3352–7.
25 Joyce JA, Pollard JW. Microenvironmental regulation of metastasis. Nat Rev Cancer.
2009;9:239–52.
26 Chiang AC, Massague J. Molecular basis of metastasis. N Engl J Med. 2008;359:2814–23.
27 Kansas GS. Selectins and their ligands: current concepts and controversies. Blood.
1996;88:3259–87.
28 Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the
leukocyte adhesion cascade updated. Nat Rev Immunol. 2007;7:678–89.
29 Polgar J, Matuskova J, Wagner DD. The P-selectin, tissue factor, coagulation triad. J Thromb
Haemost. 2005;3:1590–6.
30 Borsig L, Wong R, Hynes RO, Varki NM, Varki A. Synergistic effects of L- and P-selectin in
facilitating tumor metastasis can involve non-mucin ligands and implicate leukocytes as
enhancers of metastasis. Proc Natl Acad Sci. U S A 2002;99:2193–8.
31 Laubli H, Spanaus KS, Borsig L. Selectin-mediated activation of endothelial cells induces
expression of CCL5 and promotes metastasis through recruitment of monocytes. Blood. 2009.
32 Laubli H, Stevenson JL, Varki A, Varki NM, Borsig L. L-selectin facilitation of metastasis
involves temporal induction of fut7-dependent ligands at sites of tumor cell arrest. Cancer Res.
2006;66:1536–42.
33 Psaila B, Lyden D. The metastatic niche: adapting the foreign soil. Nat Rev Cancer. 2009;9:285–
93.
34 Laubli H, Borsig L. Selectins promote tumor metastasis. Semin Cancer Biol. 20:169–77.
35 Stevenson JL, Choi SH, Varki A. Differential metastasis inhibition by clinically relevant levels of
heparins – correlation with selectin inhibition, not antithrombotic activity. Clin Cancer Res.
2005;11:7003–11.
36 Ludwig RJ, Alban S, Bistrian R, Boehncke WH, Kaufmann R, Henschler R, Gille J. The ability
of different forms of heparins to suppress P-selectin function in vitro correlates to their inhibitory
capacity on bloodborne metastasis in vivo. Thromb Haemost. 2006;95:535–40.
37 Hostettler N, Naggi A, Torri G, Casu B, Vlodavsky I, Borsig L. P-selectin- and heparanase-
dependent antimetastatic activity of non-anticoagulant heparins. FASEB J. 2007;21:3562–72.
2
1
/
5
100%