voies visuelles, vision binoculaire et oculomotricite plan

1
Ophtalmologie, cours n° 7
Le 11 décembre 2008, 8 h 30
Professeur : Emmanuel Bui Quoc
Ronéotypeur : Ferdi Kundul
VOIES VISUELLES, VISION BINOCULAIRE ET
OCULOMOTRICITE

2
PLAN
Introduction
I – LES VOIES VISUELLES
1) La mécanique visuelle – la rétinotopie
2) Les voies visuelles de l’œil au cortex strié (ou cortex visuel primaire)
3) Le message visuel
4) L’organisation anatomique du corps géniculé latéral dorsal
5) L’organisation fonctionnelle du cortex visuel primaire
5.1) les colonnes de dominance oculaire
5.2) Les colonnes d’orientation
5.3) Les « blobs »
5.4) Les cellules binoculaires
5.5) Les hypercolonnes
6) Les cortex extra- striés (ou cortex visuels secondaires)
7) Physiologie du corps calleux
8) Déficits du champ visuel
II – VISION BINOCULAIRE
1) Exemple d’un sujet adulte atteint de diplopie bincolaire
2) Exemple d’un enfant atteint de strabisme
3) Différence entre strabisme et diplopie
III – OCULOMOTRICITE
1) Loi de Hering
2) Exemple d’un sujet adulte atteint de paralysie oculomotrice
3) Exemple d’un enfant atteint de paralysie oculomotrice

3
Introduction
Ce cours à pour principal objectif de montrer que le système visuel est un système qui est, certes,
sensoriel mais aussi oculomoteur puisqu’il est nécessaire d’avoir une oculomotricité correcte de telle sorte
que l’image soit sur les fovéas afin d’avoir une bonne perception. Nous nous pencherons sur des
altérations de la fonction visuelle telles que la diplopie et le strabisme de l’enfant. Soulignons dès à
présent que strabisme n’est pas égal à diplopie et que devant un tableau de sujet qui louche il faut faire la
différence entre paralysie et strabisme parce que les conséquences thérapeutiques et diagnostiques sont
différentes.
L’œil est un appareil qui permet de focaliser l’image sur la rétine. Il y a, certes, une intégration du
message rétinien au niveau de l’œil, mais la perception visuelle est une perception corticale : c’ est
pourquoi nous étudierons les voies visuelles. Les éléments majeurs de la perception sont les cortex
occipitaux mais dans la perception visuelle sont impliquées d’autres éléments tels que les aires
associatives et le corps calleux. Quant aux effecteurs moteurs, ils permettent de viser la cible.
L’acuité visuelle telle qu’on la mesure en pratique est une mesure psycho-physique dans laquelle on
détermine l’angle minimum de résolution entre deux lignes. Une acuité visuelle de 10/10 correspond à la
perception d’1 minute d’arc. L’acuité visuelle se mesure à différents contrastes. Les optotypes en noir sur
blanc ont un contraste égal à 1 : c’est la mesure de la meilleure acuité visuelle. Celle-ci est moins bonne
lorsque le contraste diminue, i.e. lorsque les tons deviennent plus gris. L’acuité visuelle est aussi la vision
des couleurs et la vision stéréoscopique. Même sur une photographie plane on a une idée du relief parce
que, comme nous le verrons, la vision stéréoscopique ne procède pas uniquement de la disparité d’image
entre les deux images qui se forment sur les deux rétines : d’autres éléments nous permettent d’affirmer
qu’un objet est plus près ou plus loin qu’un autre.
La vision est donc une fonction complexe dont la connaissance de la physiologie est favorisée par la
facilité d’accès du récepteur (l’œil) et de l’intégrateur principal (cortex occipital). Nos connaissances sur
le système visuel chez l’humain sont essentiellement fondées sur des observations neuropsychologiques
(observations sur des patients présentant des lésions focales du système visuel) et sur des extrapolations
d’observations faites en électrophysiologie chez l’animal.
La cascade « stimulus→information→récepteur→transmetteur », a lieu au niveau rétinien. Ensuite la
voie dite « décodeur→ intégrateur » correspond aux voies visuelles, et à l’intégration du message au
niveau cortical.
I – LES VOIES VISUELLES
1) La mécanique visuelle – la rétinotopie
Les voies visuelles comprennent :
le corps géniculé latéral dorsal (CGLd)
le cortex visuel primaire
les cortex visuels secondaires
le corps calleux

4
La rétine est d’abord un récepteur d’image de l’espace objet qui, en dehors de la rétine centrale (fovéa), s’organise
en 3 étages :
1er étage : les photorécepteurs,
2ème étage : les cellules amacrines (ou cellules bipolaires),
3ème étage : les cellules ganglionnaires
On a au niveau rétinien toute une machinerie cellulaire qui va déjà être l’intégration du message : 100
millions de photorécepteurs au niveau de la rétine transmettent le message visuel à seulement 1 million de
fibres au niveau du nerf optique. L’image (information sensorielle) sur la rétine, si on l’assimile à un
écran avec des pixels, c’est un pixel qui donne une réponse au bout d’un neurone au niveau cortical. Il y a
une rétinotopie : des zones du cortex occipital correspondent à certaines régions de la rétine mais il y a
une intégration de ce message puisqu’ il n’y a pas « 1 point pour 1 point » (100 millions de
photorécepteurs et uniquement 1 million de fibres optiques).
2) Les voies visuelles de l’ œil au cortex strié (ou cortex visuel primaire)
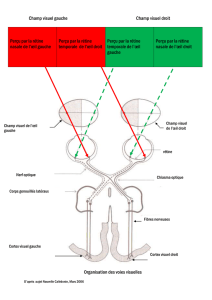
Voies de conduction visuelles :
Deux nerfs optiques
↓
Chiasma
↓
Tractus optique
↓
Corps géniculé latéral dorsal
↓
Radiations optiques
↓
Cortex visuel primaire (ou cortex strié ou cortex V1) et aires associatives
On remarque le corps géniculé latéral dorsal situé au centre des visuelles.

5
Il y a une organisation particulière des fibres des cellules ganglionnaires (3ème étage de la rétine) qui vont
des différentes zones du champ rétinien vers le nerf optique, une organisation aussi de ces fibres
nerveuses au niveau du nerf optique et, ensuite, au niveau des voies réticulées statiques. Ceci va
permettre, quand on analyse un déficit du champ visuel, de déterminer la localisation de la lésion, qu’elle
soit rétinienne ou au niveau du nerf optique.
3) Le message visuel
Le message visuel arrivant à la rétine est une radiation électromagnétique (lumière), un stimulus dont les
différentes caractéristiques sont : la longueur d’onde en fonction de laquelle on a une perception colorée
différente, l’intensité lumineuse, et aussi un contraste et un mouvement.
Rappelons qu’il y a une décussation d’une partie des fibres optiques puisque les fibres qui viennent de la
rétine temporale sont directes alors que les fibres qui viennent de la rétine nasale sont croisées. Quant aux
fibres qui correspondent aux 2 degrés du méridien vertical central, elles ont une double représentation :
une dans chaque hémisphère puisqu’elles sont à la fois directes et croisées. De plus cette zone centrale
correspondant à la macula est reliée à l’autre hémisphère par des connections inter-hémisphériques via le
corps calleux.
Tout au long des voies visuelles, l’information passe de l’œil au corps géniculé latéral, puis au cortex
occipital : en fait, c’est un système uniquement à 3 neurones parce que la cellule ganglionnaire donne un
relais synaptique au niveau du corps géniculé latéral dorsal à un autre neurone qui envoie à son tour son
information au niveau des cortex occipitaux.
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%