Cours NBC 2015_complet_complet [Mode de compatibilité]

Neurobiologie de la mémoire
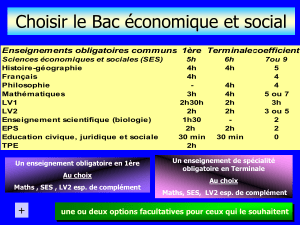
HMBS106
HMBS106
Michel Vignes

•Recherche des mécanismes moléculaires et cellulaires responsables
de la mémorisation: qu’est-ce qui se passe dans un neurone lorsqu’il
y a une mémorisation? Où sont stockées les informations dans le
système nerveux central?
•Interférence avec les processus de mémorisation: maintien de
certaines informations et oubli d’autres (mémoires aversives);
La mémoire et les neurobiologistes
certaines informations et oubli d’autres (mémoires aversives);
•Utilisation de modèles animaux ‘simples’ comportant des circuits
synaptiques facilement identifiables et des comportements
stéréotypés pouvant être corrélés avec une activité synaptique;

Aplysie
Drosophile
Abeille
Souris
Rat Humain
Diamant mandarin

Les différentes formes de mémoire humaine
•De manière intuitive, on peut classer la
mémoire en : à court terme et à long terme.
–Je me souviens d’un code ou d’un n° de téléphone
qu’on me dit avant de le chiffrer sur un clavier
qu’on me dit avant de le chiffrer sur un clavier
(court terme);
–J’ai appris à faire du vélo et je m’en souviendrai
toute ma vie.

Les différentes formes de mémoire humaine
•On peut aussi distinguer différentes formes de mémoire: consciente et
inconsciente
–Mémoire ‘consciente’ ou déclarative :
•Mémoire sémantique : je me souviens du langage, des mots, des idées, du ‘savoir universel’
comme ‘Paris est la capitale de la France’;
•Mémoire épisodique : je me souviens des évènements de ma vie et du monde qui m’entoure, je
suis capable de faire des nouveaux souvenirs. Cette forme de mémoire s’altère avec le
vieillissement et les pathologies neurodégénératives comme la maladie d’Alzheimer;
–
Mémoire ‘inconsciente’:
–
Mémoire ‘inconsciente’:
•Mémoire ‘procédurale’: j’ai appris à faire du vélo, à conduire une voiture et je m’en souviens de
manière inconsciente chaque fois que je suis sur un vélo ou au volant d’une voiture;
•Mémoire ‘émotive’ : elle fait appel à des peurs inconscientes. Serait à l’origine des ‘phobies’;
•Apprentissage associatif: deux évènements non reliés entre eux vont être associés. On parle aussi
de conditionnement ou apprentissage de type ‘pavlovien’. On associe facilement une personne et
un parfum par exemple.
•Apprentissage non-associatif:
–Habituation: on répond de moins en moins à des stimuli s’ils sont répétés (et inoffensifs); Par exemple on
ne réagit plus face à un environnement connu (rue, domicile);
–Sensibilisation : on de vient plus sensible à des stimulations qui ne nous font rien habituellement: on
devient plus attentifs aux sons qui nous entourent dans l’obscurité.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
1
/
150
100%