le cytosquelette

UE2 –Séance 3

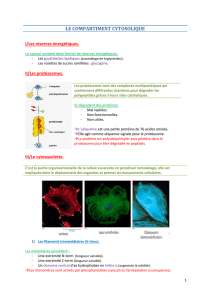
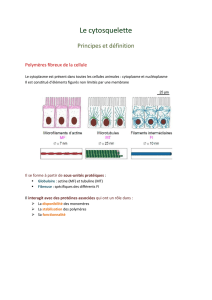
Généralités
Polymères peu mobiles formés à partir de petites
molècules très mobiles dans la cellule
Un polymère est formé de plusieurs protofilaments
(mise bout à bout de monomères)
Très résistants, très dynamiques (->adaptation de la
cellule)


Les Microtubules
Formé de 13 protofilaments de 5 nm de diam
Structure cylindrique creuse
Diamètre = 25 nm
Tubuline = prot globulaire (55kDa)
7 types de globuline (α, β, γ+++)
Hétérodimère αβ = SU du MT, lié au GTP
Seul le GTP de la tubuline β s’hydrolyse en GDP
(stabilité des dimères)

MT labiles = cytosoliques, labiles à basse t° (4°C),
sensibles aux alcaloïdes dépolym (colchicine…)
MT stables = résistants+++, structures pluritubulaires
(centrioles, corpuscules basaux, cils, flagelles)
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
1
/
40
100%