La contraction musculaire normale L

Les muscles se raccourcissent sur commande. Ils ont une architecture hypersophistiqués.
Les muscles représentent 30 à 40% du poids du corps. Ils sont composés de 75 à 80% d'eau, 15 à 20% de
protéines, 1% de glycogène, 1% de lipides, 1% de sels minéraux. Il existe deux sortes de muscles : les muscles
striés (muscles squelettiques et myocarde) et les muscles lisses.
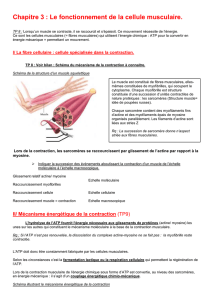
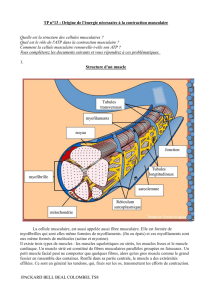
Le muscle squelettique - dont la fonction est de mobiliser les différentes parties du squelette - est
un ensemble de cellules (fibres musculaires) qui forment l'unité fonctionnelle du muscle. Leur diamètre varie de
0,01 à 0,1 mm et leur longueur peut atteindre 10 cm. Elles se rassemblent en faisceaux qui fusionnent avec les
fibres tendineuses.
Les fibres musculaires sont composées
d'une membrane (le sarcolemme)
d'un cytoplasme (le sarcoplasme)
de plusieurs centaines de noyaux rejetés à la périphérie
d'organismes intra cytoplasmiques (très nombreuses mitochondries, réticulum sarcoplasmique en
réseau...)
de myofibrilles organisées en unités fondamentales, les sarcomères, reliées entre elles pour former un
cytosquelette, lui-même fixé au sarcolemme. Les principales myofibrilles sont : la Myosine et le
complexe actinique (Actine, Troponines, Tropomyosine, ...)
de myoglobine, d'enzymes...
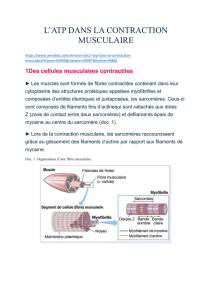
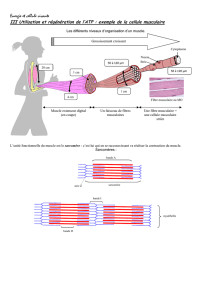
Vue microscopique d'un muscle
squelettique ; on distingue bien les
stries caractéristiques qui
correspondent à la juxtaposition des
sarcomères
Schéma d'un sarcomère : les stries Z,
sur lesquelles sont fixés les complexes
actiniques, sont reliées entre elles et à
la membrane cellulaire. La disposition
imbriquée des filaments d'Actine et de
Myosine permet leur glissement
réciproque.
La contraction musculaire normale

L'excitation du moto neurone induit une libération d'acétylcholine dans la fente
synaptique de la plaque motrice ; l'acétylcholine se fixe sur son récepteur, active
simultanément les canaux à sodium et à potassium. L'irruption brutale du Na+ dans la cellule
associée à la sortie brutale de K+ crée un potentiel électrique appelé onde de dépolarisation.
Par activation successive des canaux sodiques et potassiques, la dépolarisation
se propage le long de la membrane plasmique qui est invaginée à l'intérieur des cellules
musculaires formant un réseau de canaux intra-cellulaires (les tubules transverses en T). Il y a
2 invaginations par sarcomère. La dépolarisation se propage ainsi à l'intérieur de la cellule,
atteignant simultanément et "immédiatement" l'ensemble des myofibrilles.
Les tubules en T sont en connexion avec les réservoirs sarcoplasmiques
contenant le Ca++ fixé à une protéine, la calséquestrine. Quand la dépolarisation atteint ces
réservoirs, ils libèrent très rapidement (1 à 2 milliseconde) une grande quantité de Ca++ dont
la concentration intracellulaire est multipliée par 100 à 1000.
Le Ca++ se fixe sur la Troponine C (qui possède 4 sites de fixation), entraînant
le changement de conformation de la tropomyosine disposant ainsi les sites actifs de l'actine
en face de ceux de la myosine.
En présence d'ATP, il y a formation d'un complexe à haute énergie Myosine-
ADP-P ; celui-ci se fixe sur l'actine, libère ADP + P, récupérant ainsi l'énergie nécessaire à la
modification de l'angle de la tête de la myosine ; en pivotant de 45°, cette tête déplace le
filament d'actine de 10 nm en direction du centre du sarcomère, raccourcissement d'autant la
longueur de la fibre. La force engendrée par cette modification de structure de la myosine est
de l'ordre de 5 à 10-12 Newton.
En présence d'une seconde molécule d'ATP, il y a formation d'un complexe
Actine-Myosine-ATP qui a une faible affinité pour l'actine ; il y a dissociation de la liaison
Actine-Myosine avec libération d'ADP + P. Le tout (formation du complexe Myosine-ADP-P,
formation du pont Actine-Myosine, déplacement, rupture du pont Actine-Myosine) ne dure
que quelques millisecondes.
A ce moment là, il y a deux possibilités :
soit d'autres fibres musculaires ont aussi été excitées, entraînant un raccourcissement
du muscle ; dans ce cas, les 10 nm de raccourcissement ne sont pas perdus et, si la
fibre reçoit un nouveau potentiel de plaque motrice, le même cycle permettra de
gagner 10 nm de plus ; le filament d'Actine progresse ainsi de 10 nm en 10 nm le long
du filament de Myosine, comme une chaîne sur une roue dentée. Le processus
s'interrompt quand la tête de la Myosine atteint la dernière molécule d'Actine sur le
filament fin.
soit les autres fibres n'ont pas été excitées, le muscle se relâche et, s'il est soumis à un
étirement, les filaments d'Actine et de Myosine glissent l'un sur l'autre pendant que le
muscle s'allonge.

La mise en jeu de plusieurs millions de liaisons Actine-Myosine induit une
force de plusieurs millions de fois 5 à 10-12 Newton mais aussi une consommation de
plusieurs millions de molécules d'ATP. La succession de plusieurs cycles d'excitation par
seconde (40 à 100) produit le raccourcissement du muscle.
La mise en route de la contraction musculaire est donc une dépolarisation
électrique, mais le moteur énergétique est la consommation de molécules d'ATP. Quels sont
les mécanismes produisant cette molécule ?
Ils sont de deux sortes :
les mécanismes extrêmement rapides, pouvant répondre à une demande de plusieurs
millions de molécules en un temps très court (permettant ainsi les contractions
musculaires rapides) :
* la phosphorylation directe d'une molécule d'ADP selon la réaction ADP + P
==> ATP
* la glycolyse intra cytoplasmique anaérobie selon la réaction : 1 glucose ==> 3
ATP + 2 Pyruvate ; le pyruvate formé sera ensuite brûlé dans le cycle de Krebs
intramitochondrial.
les mécanismes plus lents correspondants à la combustion aérobie intramitochondriale,
dont le rendement est nettement meilleur :
* des pyruvates : 35 ATP pour les 2 pyruvates issus d'une molécule de glucose
* des acides gras : 128 ATP pour la combustion d'une molécule de palmitate
(acide gras à 16 atomes de carbone).
* des corps cétoniques : 26 ATP pour la combustion d'un acide bêta hydroxy-
butyrique
Les sous-produits de cette combustion aérobie sont : le CO2, qui devra être
éliminé par les poumons, l'eau et la chaleur.
La relaxation musculaire enclenchée, le calcium sera transféré dans les
réservoirs sarcoplasmiques au moyen d'une pompe (fonctionnant à l'ATP) et le muscle
reviendra à son état de relâchement.
1
/
3
100%