LA SPERMATOGENESE

MODULE de MAIEUTIQUE
Anatomie et histologie de l’appareil reproducteur et du sein, organogénèse,
tératogenèse
Histo-Embryologie
(Coordonateur Pr Janky)
Histologie des gonades, du sein et de l’appareil
génital
Dr. Nadia Prisant

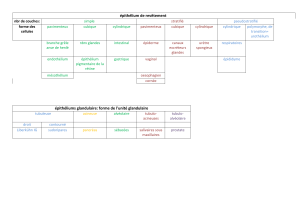
GONADES
TESTICULES
Les testicules humains sont deux organes de forme ovoïde, dont les dimensions moyennes sont
d’environ 5 cm de long sur 3 cm de large et 2,5 cm d’épaisseur.
Le testicule a une fonction exocrine et une fonction endocrine. La fonction exocrine est la production
des gamètes contrôlée par les cellules de Sertoli qui libèrent dans la lumière des tubes séminifères,
les spermatozoïdes à partir des cellules germinales.
La fonction endocrine est assurée par les cellules interstitielles de Leydig
Il existe un important contrôle paracrine local.
Les cellules de Sertoli
Elles représentent 17% du volume testiculaire. Ce sont des cellules somatiques qui ont un rôle de
soutien pour les cellules germinales. Elles sont indispensables au bon déroulement de la
spermatogenèse. Leur différenciation débute peu avant la puberté.
Les cellules de Sertoli sont des cellules pyramidales, de grande taille (70 x 20 µm), qui reposent sur la
membrane basale. Leur pointe atteint la lumière du tube. Les limites cellulaires ne sont pas visibles
en microscopie photonique (aspect pseudo-syncitial). Seule la microscopie électronique a permis de
préciser leur morphologie.
- Leur contour est irrégulier et les faces latérales sont
déprimées par les cellules germinales voisines.
- Le cytoplasme, pâle, renferme de volumineuses enclaves
lipidiques, associées à du réticulum lisse, de nombreux
lysosomes (primaires et secondaires) et des pigments
(lipochromes). Il renferme des inclusions caractéristiques, les
cristalloïdes de Charcot (ou Charcot-Bottchner). Elles
mesurent 2 à 4 µm et sont constituées des microtubules
rangés parallèlement. On y décrit également les cristalloïdes
de Lubarsch, allongés, et les cristalloïdes de Spangaro,
colorables seulement par certaines méthodes.
- Le noyau, au tiers basal, triangulaire ou ovalaire, a un
grand axe perpendiculaire à la basale. Il est souvent encoché
et est nucléolé (le nucléole est caractéristique, avec une
masse centrale acidophile encadrée par 2 blocs basophiles).
La barrière hémato-testiculaire
Les cellules de Sertoli sont réunies entre elles par des complexes de jonction au pôle basal, par
quelques jonctions communicantes et par des desmosomes sur leurs faces latérales. En regard des
cellules germinales, les espaces intercellulaires sont assez larges (15 à 20 nm) et la membrane
plasmique ne présente pas de différenciation notable, sauf au niveau des spermatides. A ce niveau,
une couche fibrillaire sous-membranaire, riche en actine, est associée à des citernes lamellaires de
réticulum lisse. Au tiers externe de l'épithélium, des jonctions serrées (Zonula occludens) (ZO) entre
les cellules de Sertoli séparent l'épithélium séminal en 2 compartiments :
- Un compartiment externe où se trouvent les spermatogonies et spermatocytes jusqu'au
stade préleptotène.

- Un compartiment interne, communiquant avec la lumière du tube, qui renferme les
éléments les plus avancés de la lignée germinale. Les cellules de Sertoli sont les seuls
intermédiaires entre le sang et cet espace interne.
Les cellules de Leydig
Elles représentent environ 3% du volume testiculaire et constituent la glande interstitielle du
testicule.
Morphologie
Ce sont des éléments polyédriques de 15 à 20 µm de diamètre,
au noyau rond central, nucléolé et à la chromatine granulaire.
Le cytoplasme est comparable à celui des autres cellules
élaborant des hormones stéroïdes. Il est dense ou
microvacuolaire, riche en réticulum lisse, en mitochondries (à
crêtes tubulaires), en vitamine C et en enzymes de la synthèse
des stéroïdes. Il renferme des vacuoles lipidiques riches en
cholestérol et des enclaves pigmentaires. Le cytoplasme
renferme en outre des inclusions protéiques à structure
cristalline : les cristalloïdes de Reinke.
Biologie
La cellule de Leydig élabore les androgènes testiculaires (Testostérone, androstènedione, Déhydro-
Epi-Androstérone, Dihydrotestostérone, androstanediol). Ces androgènes sont synthétisés à partir du
cholestérol. L'androgène principal chez l'Homme est la testostérone qui, en synergie avec la FSH,
assure le maintien et l'intégrité de la lignée germinale. Elle agit également sur d'autres tissus : tissu
osseux, tissu musculaire, peau et ses annexes. La cellule de Leydig libère également un peu
d'estrogènes, qui, avec les estrogènes libérés par les cellules de Sertoli totalisent 10 à 20 % des
estrogènes chez l'homme. L'activité de la cellule de Leydig est stimulée par la lutéotropine
hypophysaire (L.H.), produite par les cellules gonadotropes de l'hypophyse glandulaire. La cellule de
Leydig est sensible à d'autres médiateurs (prolactine, glucocorticoïdes, insuline, EGF, vitamines A et
E, prostaglandines). Inversement, les stéroïdes sexuels, androgènes et estrogènes exercent un effet
inhibiteur sur l'activité de la cellule de Leydig. Ainsi il existe un rétrocontrôle négatif de la production
des stéroïdes sexuels à l'intérieur même du lobule testiculaire.
La spermatogénèse
La spermatogenèse est l’ensemble des divisions cellulaires et des différenciations au cours de
l’évolution de la lignée germinale qui, à partir des cellules souches (spermatogonies), aboutissent à la
formation des gamètes mâles matures (spermatozoïdes). C’est un processus continu dans le temps à
partir de la puberté.
Dans, la lignée germinale, 3 types de cellules sont reconnaissables, correspondant chacun à une
étape précise:
o les spermatogonies: divisions mitotiques
o les spermatocytes: méiose
o les spermatides: différenciation terminale ou spermiogenèse

La durée complète de la spermatogenèse humaine, appelée cycle spermatogénétique, est de 74
jours. La méiose spermatocytaire débute à la puberté, immédiatement après une phase de
multiplication des spermatogonies. Ces spermatogonies se sont développées à partir des cellules
germinales primordiales qui ont colonisé le testicule fœtal
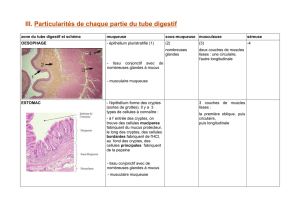
Le cycle de l’épithélium séminal
L’observation, en coupe transversale, des tubes séminifères montre qu’ils n’ont pas tous le même
aspect, c’est à dire qu’ils n’ont pas la même composition en cellules germinales de différents types
Chez l’homme, on peut observer 6 associations préférentielles de cellules germinales entre elles
définissant les 6 stades de l’épithélium séminal
Ces stades sont dus à une entrée en mitose cyclique et régulière des spermatogonies associée à une
durée d’existence variable de chaque type cellulaire pendant la spermatogenèse
Les spermatogonies entrent en division de façon périodique, tous les 16 jours.
Parallèlement à ce phénomène, chaque type de cellule germinale a une durée de vie différente.
Les spermatozoïdes libérés par l’épithélium séminal sont transportés le long de l’épididyme pendant
12 jours supplémentaires. Ainsi, le cycle complet de la spermatogonie jusqu’au spermatozoïde
mature dure 86 jours.
18j 9j
23j
1j
23j
Chacun des 6 stades se répète tous les 16j = durée du cycle de l’épithélium séminal
74j
Cycle spermatogénétique = 74j ~ 4 cycles et demi de l’épithélium séminal

OVAIRES
Les fonctions de l'ovaire sont l'ovogenèse et la stéroïdogenèse. L'œstrogène et la progestérone
préparent l'endomètre de l'utérus pour l'implantation et le maintiennent en cas de fécondation.
La surface de l'ovaire est couverte d'un épithélium simple (cubique ou pavimenteux) continue avec le
mésothélium du péritoine
La tunique albuginée est une couche de tissu conjonctif dense,au-dessous de l'épithélium.
Le Cortex ovarien est formé de 2 composantes:
1. Stroma cortical : (tissu de soutien) formé des fibres du tissu conjonctif et d'un grand nombre
de cellules du stroma
2. Les follicules ovariens formés d'ovocytes et de cellules folliculaires. Ces follicules sont dans
différents stages.
La médullaire est formée de tissu fibro-élastique lâche, quelques muscles lisses, des vaisseaux
sanguins et lymphatiques ainsi que des nerfs Les vaisseaux et les nerfs rejoignent la médullaire à
travers le hile.
Ovogenèse
1. Les ovogonies sont les cellules germinatives. Chez le fœtus féminin, au cours du 2e trimestre
de la grossesse, toutes les ovogonies augmentent de taille et se nomment ovocytes de 1er
ordre (ovocytes I).
2. Les ovocytes I commencent la prophase de la 1ère division méiotique; ils restent bloqués à
cette phase jusqu'à ce que les cycles ovariens et menstruels commencent à la puberté.
3. À chaque cycle quelques-uns des ovocytes I continuent à se développer. Juste avant
l'ovulation, l'ovocyte I complète la 1ère division méiotique. Résultats : formation de l'ovocyte
du 2e ordre (ou ovocyte II) ainsi qu'un 1er globule polaire (ce dernier contient l'excès du
matériel génétique).
N.B. Les ovogonies et les ovocytes I contiennent 46 chromosomes "2n" (22 autosomes et 2 sexes
chromosomes "XX" et sont des cellules diploïdes, tandis que l'ovocyte II est une cellule haploïde
i.e. 23 chromosomes "n" (22 autosomes et 1 sexe chromosome "X")
4. L'ovocyte II commence immédiatement la 2edivision méiotique, il s'arrête à la métaphase et
ne se complète qu'en cas de fertilisation.
Résultat : ovule + un 2e globule polaire. Le 1er globule polaire ne se divise pas, les globules
polaires dégénèrent.
N.B. L'ovogonie donne 1 ovule, tandis que la spermatogonie donne 4 spermatozoïdes.
Développement des follicules ovariens
Dans la partie externe du cortex ovarien, se trouve un grand nombre (600 000 - 800 000) de follicules
primordiaux.
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%