Biologie du développement végétal

Biologie de la reproduction et du développement végétal
Cours de E. Boncompagni
Schémas complémentaires disponibles sur
http://www.unice.fr/IPMSV/Enseignement/Boncompagni.htm
Introduction :
Dans ce cours nous traiterons uniquement des Spermatophytes (= végétaux à graine).
L’apparition de la graine est une étape importante dans l’histoire de l’évolution des plantes. On retrouve dans ces
graines tous les éléments qui ont été mis en place par les Bryophytes (mousses) et les Ptéridophytes (fougères).
Mais ici le moyen de reproduction ne sera plus la spore, elle se fera grace au gamétophyte. Ce sera le
gamétophyte femelle qui permettra la formation de l’embryon.
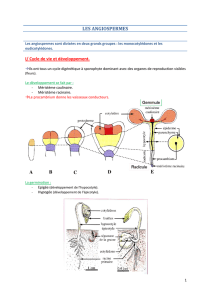
Parmi les Spermatophytes se trouvent :
- Les Gymnospermes (=graines nues) qui sont actuellement divisés en - Pinophytes
- Ginkgophytes
- Cycadophytes
- Gnétophytes
- Les Angiospermes, plantes à double fécondation, qui va permettre de produire un tissus nourricier qui aura
pour rôle de protéger la graine à l’intérieur de l’ovaire. Ce tissus va donner le fruit.
Les Angiospermes est un groupe très vaste, divisé en - Eudicotylées
- Monocotylédones
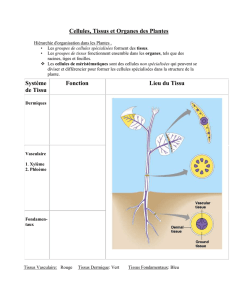
Ontogenèse végétale = Développement progressif d’un être vivant de sa conception jusqu’à sa mort.
La plante va se construire tout au long de sa vie à partir de tissus dits indifférenciés (méristèmatiques) qui
trouvent leur origine dans les méristèmes, formés de cellules indifférenciées, totipotentes, à partir desquelles les
plantes vont former les différentes lignées cellulaires.
La phase de développement embryonnaire est très réduite. La phase post embryonnaire est complète, car elle va
mettre en place les différents tissus et organes.
Avantage : le développement est adaptable et flexible selon les conditions environnementales. Les méristèmes
fonctionnent en continu : organogenèse permanente.
Deux processus distincts de développement:
- Primaire : processus mis en place très tôt au niveau de la gemmule et de la
radicule.
Il se fait grâce au méristème apical selon deux axes :
Axe caulinaire pour la partie aérienne et axe racinaire pour la partie terrestre.
Ce fonctionnement permet l’édification de l’architecture de la plante. Il existe
des massifs cellulaires conduisant à la formation de tissu primaire. Les
divisions sont très actives, et se font par de nombreuses mitoses transversales.
Ces méristèmes sont caractérisés par la mise en place de cellules désorganisées
qui vont édifier le végétal à partir d’un point d’origine méristèmatique.
- Secondaire : processus apparaissant beaucoup plus tard, au cours du
développement du végétal.
Il va servir principalement à la croissance en épaisseur de la tige et de la racine,
qu’on retrouve dans les plantes ligneuses. Ces méristèmes sont caractérisés par
la mise en place de structures présentant des files cellulaires qui vont rayonner
de part et d’autre d’une assise. Activés, ces méristèmes vont avoir des rythmes de fonctionnement bien
particuliers, en correspondance avec l’alternance des saisons.
Deux types de méristèmes secondaires :
- Assise libéro-ligneuse liber + bois
- Assise subéro-phellodermique suber + phelloderme

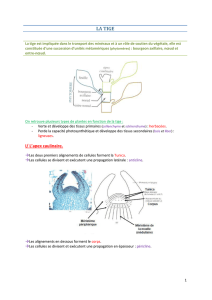
La Racine
Organe généralement sous-terrain. Rôle essentiel dans l’absorbtion de l’eau et des sels minéraux, et dans la
fixation de la plante au sol. La racine présente un gravitropisme positif. Elle a un rôle de stockage de différentes
substances de réserve et dans la synthèse de différents composés comme les hormones végétales (cytokinine,
acide abscissique (ABA)). Ils sont produits au niveau de la racine, puis migrent dans le reste de la plante.
I) Structure externe
Au moment de la germination, la radicule sort et s’allonge pour
former la racine, organe cylindrique de symétrie axiale.
Il n’y a ni feuilles ni structure définie.
Il existe plusieurs zones :
- La Coiffe, en amont de la racine
- La Zone lisse
- La Zone pilifère portant les poils absorbants
- La Zone subéreuse, ébauche de la racine secondaire.
II) Point végétatif
A) Organisation de l’extrémité de la racine
L’extrémité de la racine est recouverte d’un tissu fragile : la coiffe. Elle a un rôle de protection du méristème.
Ses cellules vont libérer un mucilage (composés organiques lubrifiants) qui va faciliter la croissance de la racine
dans le sol.
La racine est capable de percevoir la gravité grâce à des cellules de la coiffe qui jouent le rôle d’organes
sensoriels : les statholites (grains d’amidon mobiles). Ils vont orienter la plante.
Les cellules de la zone quiescente, au centre, sont une zone méristématque très
faible. C’est le point de développement du méristème, il y a très peu de divisions
cellulaires.
Par contre, la zone active est le lieu de divisions intenses.
Le méristème apical racinaire va être composé de différents tyes de tissus qui vont se multiplier :
1) Dans la partie centrale : Procambium = permet les divisions cellulaires de la partie centrale.
2) De chaque côté : Méristème fondamental.
3) Au bord : Protoderme.
Ces différentes cellules mettent en place différents tissus racinaires : dans la zone d’élongation, la plupart des
tissus primaires vont se différencier. Mise en place des phloèmes et xylèmes (vaisseaux).

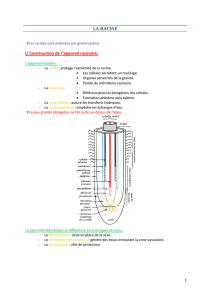
III) Structure primaire
A) Coupe transversale
Chez les Eudicotylédones :
Zone fonctionelle différenciée :
Assise formée par une couche continue de cellules aux membranes minces. Chaque cellule est
connectée avec la cellule mitoyenne. Certaines cellules vont emettre des poils absorbants qui vont
assurer un rôle de transport de l’eau du sol vers le centre de la structure racinaire. Tissu à origine
méristèmatique. Il vient du protoderme.
Exoderme situé sous l’assise pilifère. Il est formé de cellules vivantes disposées sur une ou plusieurs
couches.
1 seule couche = assise.
Plusieurs couches = zone cellulaire.
Paroi rapidement subérifiée dès la perte des poils absorbants. Zone hydrophobe empêchant les échanges
d’eau avec l’extérieur.
Cortex (parenchyme) : tissu constitué de cellules aux parois cellulosiques. Entre les cellules se trouvent
les méats qui laissent passer l’eau. Rôles de cellules : stockage des sels minéraux et de l’eau. Zone
facilitant les transferts. Lors du vieillissement, les cellules s’épaississent et leurs parois se sclérifient.
Tissu vasculaire :
Endoderme situé vers la stelle. Parois très épaisses. L’endoderme est formé de cellules qui limitent
intérieurement l’écorce, caractéristiques par la constitution de leurs membranes. Les membranes
latérales de chaque cellule portent des épaississements en libéro-lignine.
L’endoderme va jouer le rôle de filtre pour les nutriments. Ainsi toutes les substances qui doivent
rentrer ou sortir du cadre central doivent passer par ces cellules.

Jusqu’ici les différentes assises se sont mises en place à partir du protoderme et du méristème fondamental.
Cylindre central (stéle) : ensemble des tissus du cylindre. Les tissus conducteurs sont localisés dans
cette partie. Ils sont séparés de l’endoderme par une première assise, le péricycle qui dérive du
procambium. Elle est formée d’une seule couche de cellules la plupart du temps, qui sont
cellulosiques. Les nouvelles structures mises en places vont se former à partir de ce tissu indifférencié.
Entre les cellules vasculaires on va trouver des cellules de parenchyme médulaire dans la jeunesse de la
plante. Ensuite elles vont disparaître.
Plaqué contre le péricycle, il y a alternance entre les pôles de xylème (ligneux ou vasculaire) et de
phloème (criblé). Ces structures se développent vers le centre de la racine : force centripète.
On a différents types de cellules :
* Protoxylème qui se différencie progressivement en
* Métaxylème : transport de la sève brute.
* Protophloème qui se différencie progressivement en
* Métaphloème : transport de la sève élaborée.
Entre les vaisseaux on trouve le parenchyme interfasciculaire.
B) Comparaison Eudicotylédones/ Monocotylées
Chez les Monocotylées la sève est beaucoup plus développée tandis que la partie corticale reste assez restreinte.
On retrouve les mêmes structures et tissus que chez les Eudicotylédones.
L’endoderme est différent : en U (ou en fer à cheval), très spécifique, qui empêche le passage des
solutés. Il existe des cellules de passage à paroi cellulosique.
Le péricycle est plus complexe : c’est une zone avec plusieurs assises, qui peut se sclérifier avec l’âge.
L’alternance phloème/xylème est >10. Il y a une grande différence de taille entre le protoxylème et le
métaxylème. L’évolution est toujours centripète.
Le parenchyme médulaire est au centre.
Le parenchyme interfasciculaire joue un rôle de remplissage dans le xylème.

Tableau de comparaison entre Eudicotylédones et Monocotylées :
Racine
Eudicotylédones
Monocotylées
Rhizoderme
Unistrate ( assise)
Parfois pluristrate ( zone)
Exoderme
Unistrate ( assise)
Pluristrate ( zone)
Endoderme
A cadre subérisé
En fer à cheval
Péricycle
Une assise cellulaire
Une ou plusieurs assises cellulaires
Cylindre central
Réduit
Plus développé
Faisceaux conducteurs
Nombre réduit (<8)
Nombreux (>10)
Métaxylème
Peu différencié par rapport au
protoxylème
Très gros faisceaux très marqués
par rapport au protoxylème
Parenchyme médulaire
Cellulosique
Souvent sclérifié
C) Comparaison Eudicotylédones/ Gymnospermes
Péricycle pluristratifié. Pas de poils absorbants. Tissus conducteurs spécifiques.
IV) Rammification de la racine
La création d’une racine secondaire se fait à l’aplomb des pôles vasculaires des cellules du péricycle, qui vont
entrer en mitose pour former un primordium radiculaire. On assiste à la formation d’un nouveau méristème
vasculaire, qui crée coiffe et autres tissus.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
1
/
26
100%