Introduction

Correction bac blanc 2 SVT avril 2012
Partie I Procréation
Introduction
La testostérone est une hormone produite par les testicules. Sa production, dès la vie
embryonnaire, a permis la mise en place des organes génitaux masculins. A partir de la puberté,
elle permettra le développement puis le maintien des caractères sexuels primaires et secondaires.
Après avoir précisé le mode de sécrétion de la de testostérone, nous verrons comment la
testostéronémie (=taux plasmatique de testostérone) est maintenue constante après la puberté
permettant alors la fonction de reproduction.
I+p
0.5
1- Mode de sécrétion :
La testostérone, hormone stéroïde, est produite par les cellules de Leydig (=cellules
interstitielles) situées entre les tubes séminifères des testicules. Cette sécrétion est
globalement stable chez l’homme. Toutefois une étude plus fine met en évidence une sécrétion
pulsatile (plusieurs pulses brefs en une journée). Les pulses ayant chez l’homme une fréquence et
une amplitude relativement constante, le taux plasmatique moyen est donc toujours proche de la
valeur dite consigne.
1
2- La commande de la sécrétion de testostérone
Les cellules interstitielles n'ont pas d'activité autonome : leur fonctionnement endocrine dépend
d'un message hormonal constitué par la concentration de LH, hormone sécrétée par
l'antéhypophyse (de façon pulsatile). La LH est donc une gonadostimuline.
La sécrétion de LH par l'hypophyse est elle-même sous la dépendance d'une neurohormone, GnRH
sécrétée de façon pulsatile par des neurones hypothalamiques.
1
0.5
3. Les variations de la commande hypothalamo-hypophysaire en fonction de la testostéronémie
L’activité du complexe hypothalamo-hypophysaire est fonction de la concentration de
testostérone. Celle-ci agit à la fois sur l'hypothalamus en diminuant les pulses de GNRH et sur
l'hypophyse en freinant la sécrétion : ils sont en effet tous les deux pourvus de récepteurs
spécifiques à la testostérone : elle exerce ainsi un rétrocontrôle négatif sur le complexe.
En cas de baisse de la testostéronémie, l'action inhibitrice exercée par la testostérone sur le
complexe hypothalamo-hypophysaire diminue; cela se traduit par une augmentation de la sécrétion
de LH. Cette augmentation induit une sécrétion plus importante de testostérone et, donc, le
rétablissement de sa concentration.
Une hausse de la concentration de testostérone entraîne des effets inverses.
1.5
1
Conclusion
Le maintien de la capacité de reproduction d'un homme est sous la dépendance d'une
concentration de testostérone à peu près constante (=paramètre réglé), assurée par le
fonctionnement du complexe hypothalamo-hypophysaire et des cellules de Leydig (=système
réglant). Tout dérèglement de cet axe gonadotrope peut entraîner une déficience de la valeur de
la testostéronémie et, par là, la stérilité.
0.5
2
Rétrocontrôle
négatif

8
Partie II Exercice 1 La convergence lithosphérique
Arguments attestant l’existence passée d’une zone de subduction dans les Alpes franco-italiennes
Si l’on fait une coupe de l’Ouest vers l’Est de la carte géologique simplifiée des Alpes franco-
italiennes, on observe, au de-là du bassin sédimentaire périphérique, une succession de roches
témoignant d’un degré de métamorphisme croissant. Ces roches, d’origine profonde, se retrouvent
en surface, suite à la collision (à l’origine de l’orogénèse alpine) et à l’érosion.
0.5
Parmi ces roches métamorphiques on observe des roches issues de la transformation de la lithosphère
océanique : ceci prouve l’existence d’un océan alpin aujourd’hui disparu.
Les ophiolites, roches de la lithosphère océanique peu transformées qui se retrouvent dans
plusieurs affleurements de la partie interne, sont très caractéristiques.
D’ouest en Est, on observe aussi :
- des schistes verts à chlorite et actinote dont le domaine de stabilité est à faible pression
(0.5GPa maxi) et une température entre 300 et 450°C.
- des schistes bleus à glaucophane: le domaine de stabilité du glaucophane se situe à une pression
supérieure à 0.5GPa et une température maxi inférieure aussi à 450°C.
- de l’éclogite dont le domaine de stabilité du minéral cité (le grenat) se situe à une pression
supérieure à 1GPa.
0.5
1.5
La succession de ces 3 roches métamorphiques (schistes verts ; schistes bleus, éclogite), formées à des
pressions croissantes, témoignent de la subduction de l’Ouest vers l’Est d’une plaque océanique sous une autre
plaque chevauchante. Cette subduction est à l’origine de la disparition de l’océan alpin.
0.5
Total
3
Partie II Exercice 2 Génétique Enseignement obligatoire
On étudie la transmission de 2 gènes chez la drosophile : un gène dirigeant la longueur des
ailes ayant 2 allèles : vg récessif et vg+ dominant ; un gène dirigeant la couleur des yeux ayant
aussi 2 allèles : pu récessif et pu+ dominant.
Prouvons que l’hypothèse « les 2 gènes sont situés sur deux chromosomes distincts est
fausse ».
Le document 1a nous renseigne sur le fait que les 2 parents sont homozygotes :♂ [vg+ ;pu+] et ♀
[vg ;pu]. Ils auront donc les allèles suivants : le ♂ : (vg+vg+ ; pu+pu+) ; la ♀: (vgvg ;pupu).
La population F1 issue de leur croisement est donc homogène : tous les individus sont
hétérozygotes : leur génotype : (vg+vg ; pu+pu) ; leur phénotype, du fait de la dominance des
allèles sera [vg+ ;pu+]
Le document 1b nous renseigne sur les résultats d’un test cross : ♀ F1 x ♂ [vgpu] (homozygotes à
allèles récessifs))
On obtient : 43.5% [vg+ ;pu+] ; 6.5% [vg+ ;pu] ; 6.5% [vg ;pu+] ; 43.5% [vg ;pu]
-Si les gènes avaient été indépendants, c'est-à-dire portés par deux paires de chromosomes
0.5

homologues, nous aurions dû avoir comme résultats à ce test-cross une équiprobabilité d’apparition
des 4 phénotypes (4x 25%) du fait de la disjonction indépendante des chromosomes homologues à
l’anaphase I de la méiose. En effet, dans ce cas, les femelles F1 devaient produire 4 types de
gamètes équiprobables : vg+pu+ ; vg+pu ; vgpu+ ; vgpu. Les mâles homozygotes produisant 100% de
gamètes vgpu (allèles récessifs). La répartition des phénotypes des individus issus du test cross
découlent de ce fait, uniquement de la répartition des gamètes femelles.
- Si les gènes étaient portés par une seule paire de chromosomes (gènes liés), les femelles F1
produiraient alors 50% de gamètes avec les allèles vg+pu+ ; 50% de gamètes avec les allèles vgpu ;
les mâles produiraient toujours 100% de gamètes vgpu. La répartition des phénotypes issus du
test-cross serait donc de 50% [vg+ ;pu+] ; 50% [vg ;pu].
Ces phénotypes correspondent aux phénotypes largement majoritaires observés pour ce
croisement.
Conclusion : Les deux gènes ne sont pas portés par deux paires de chromosomes ; ils sont
liés. On pourra écrire les génotypes des parents : ♂vg+pu+//vg+pu+ et ♀vgpu//vgpu.
Le génotype des F1 : vg+pu+//vgpu.
0.5
0.5
0.5
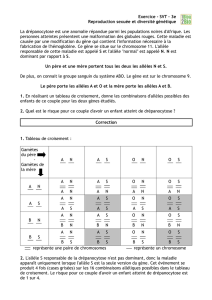
-Comment comprendre les résultats des croisements obtenus suite au test-cross ?
Le document 2 nous montre un bivalent formé par deux chromosomes homologues appariés en
prophase I de la méiose. On observe aussi que lors de cet appariement, les chromatides peuvent
se croiser (au niveau de chiasmas). Ces chiasmas peuvent s’accompagner parfois d’une rupture et
d’un échange des chromatides entre chromosomes homologues : c’est le phénomène de crossing-
over. Il y alors recombinaison génétique du fait de ce brassage intra-chromosomique.
Ce phénomène de crossing-over explique les 13% de phénotypes recombinés observés à l’issue de
ce test-cross. En effet (voir schéma d’illustration en page jointe), 13% des gamètes produits
par les femelles F1 ont hérité d’un chromosome recombiné lors de la prophase I de la méiose.
La répartition des gamètes produits par F1 est donc :
Gamètes de types parentaux :43.5% vg+pu+ ; 43.5% vgpu ; gamètes recombinés : 6.5% vg+pu ;
6.5% vg+pu
Echiquier théorique de croisement : [vg+pu+] X [vgpu] :
Gamètes ♀de
F1
Gamètes ♂
43.5%vg+pu+
43.5%vgpu
6.5%vg+pu
6.5%vgpu+
100% vgpu
43.5% vg+pu+//vgpu
[vg+ ;pu+]
43.5% vgpu//vgpu
[vg;pu]
6.5% vg+pu//vgpu
[vg+ ;pu]
6.5% vgpu+//vgpu
[vg ;pu]
L’hypothèse de deux gènes liés avec un brassage intra chromosomique dans 13% des cas
durant la prophase I de la méiose est ainsi validée
1
1
1

Total
5
16
Partie II Exercice 2 spé
Argumentation du mode autosomal récessif de la transmission de la galactosémie
-Les parents des individus malades sont sains : ils sont donc porteurs de l’allèle muté sans être
malades : l’allèle muté est donc récessif
-Les individus atteints sont des hommes et une femme : le gène ne peut donc pas être porté par le
chromosome Y
-Si l’allèle était porté par le chromosome X, le père (I2) de la femme II3 malade devrait être
malade : son chromosome X devrait être porteur de l’ alèle muté (génotype : XmY ; m : allèle
muté) :ce n’est pas le cas.
La transmission de la galactosémie s’effectue suivant un mode autosomal récessif : Allèle
N dominant normal ; m : allèle muté récessif.
1.5
Evaluation du risque pour l’enfant à naître IV-5 d’être malade.
Pour que cet enfant )à naître soit malade, il faudrait que ces deux parents soient hétérozygotes
N//m (on sait qu’ils sont sains)
Risque pour Mme G d’être hétérozygote :
Ses 2 parents sont hétérozygotes car elle a un frère (III5) malade. On sait qu’elle est saine,
suivant l’échiquier de distribution des gamètes de ses parents :
Echiquier théorique de distribution mettant en évidence la répartition de leurs descendants :
Gamètes du ♂
Gamètes de la ♀
50%N
50%m
50%N
N//N 25% [N]
N//m 25% [N]
50% m
N//m 25% [N]
m//m 25% [m]
Elle a 2/3 d’être hétérozygote.
M-G , le père n’appartient pas à cette lignée familiale, il n’a à priori aucun antécédent : son risque
d’être hétérozygote est de 1% (cf information donnée)
-Risque pour que les 2 parents soient hétérozygotes : 2/3 x 1/100= 2/300
En suivant les résultats montrés par l’échiquier précédent (avec les 2 parents hétérozygotes,
risque pour l’enfant à naître d’être malade (génotype m//m) :
2/3 x 1/100 x 1/4= 1/600
1.5

Comment le diagnostic prénatal peut-il apporter une réponse plus précise ?
-Le document 2 présente les sites de restriction de deux allèles du gène (dont 1 des deux est
l’allèle muté responsable de la galactosémie), sites de restriction résultant de l’action combinée
de deux enzymes : Seul l’allèle 2 est coupé par HpaII : Après l’action de ces deux enzymes, on
obtiendra deux fragments de restriction avec l’allèle 1 (de 111pb et de 114pb). Avec l’allèle 2, nous
en obtiendrons 3 fragments (111pb ; 73 pb ; 41 pb)
-Le document 3 présente une électrophorèse permettant de visualiser les fragments obtenus
après action sur l’ADN des différents individus de la famille
- L’homme IV4 est malade, il a donc le génotype m//m. Son électrophorèse montre que l’on a
obtenu après analyse de son ADN, les 3 fragments de l’allèle 2l’allèle 2 est donc l’allèle muté
responsable.
-L’analyse de l’ADN de Mme G fait apparaître les fragments des 2 allèles : elle est hétérozygote
N//m, comme son conjoint, M-G les deux parents sont hétérozygotes.
- L’analyse de l’ADN de l’enfant à naître IV5 ne laisse apparaître que les deux fragments
issus de l’allèle 1 L’enfant à naître est donc homozygote N//n : il est sain et non porteur.
1.5
Bilan : L’utilisation des biotechnologiques permet donc ici de lever toute ambigüité sur le
génotype des parents et de l’enfant à naître. Dans ce cas, le génotype des deux parents fera que
le risque d’avoir un enfant malade est de ¼. Ce risque important peut justifier l’utilisation de ce
diagnostic prénatal
0.5
Total
5
16
1
/
5
100%