Habilitation à diriger des Recherches Marie Bodinier 2014

Habilitation à diriger des Recherches
Université de Nantes
Ecole Doctorale VENAM
Marie Bodinier
2014
« Etudes mécanistiques du système immunitaire pour
aider à la compréhension, au diagnostic et au
traitement de certaines pathologies notamment des
allergies alimentaires. »
UR 1268 BIA
Equipe Allergie
Rue de la géraudière BP 71627, 44316 Nantes cedex 01
Illustration issue de Gurumed, 11 fév. 2013 : Des cellules de notre système immunitaire se rappellent de
microbes qu'elles n'ont jamais rencontrés auparavant

1
SOMMAIRE
DOSSIER SCIENTIFIQUE……………………………………………………………….....4
Curriculum Vitae……………………………………………………………………………..5
Liste des publications…………………………………………………………………………9
Introduction……………………………………………………………………………….…15
I. Le système immunitaire....15
A. Deux systèmes 15
B. La reconnaissance par le système immunitaire..16
C. La médiation du système immunitaire.16
D. Les lymphocytes T
1.Les différents types de lymphocytes T et leur fonction16
2.Les récepteurs des lymphocytes T.19
II. Les Allergies Alimentaires
A. Généralités sur les allergies..21
1. Prévalence.21
2. Les allergies alimentaires..22
3. 23
B. Les mécanismes de la tolérance orale et 24
1. La tolérance .24
2. La réponse allergique26
3. Modèles cellulaires et animaux.29
a. Modèles cellulaires et organes ex vivo.29
b. Modèles animaux30
III. Des stratégies nutritionnelles pour lutter contre le.31
A. La stratégie des prébiotiques, des probiotiques et des synbiotiques ...31
B. La stratégie des p
Notice des titres et travaux retraçant les résultats les plus significatifs depuis le début de
carrière………………………………….……………………………………………………34
De 1998 à 2001 : Résultats marquants au cours de mon doctorat : les mécanismes associés
aux lymphocytes T CD8 dans le cancer et les infections virales..35
I. Etude 1 : Rôle du CD8 et du TCR dans la sélection des lymphocytes T anti-viraux...35
A. But et intérêt scientifique.35
B. Programme et méthode de recherche....35
C. Résultats
II. Etude 2 -viraux...36
A. Programme et méthode de recherche....36
B. Résultats37
III. Etudes collaboratives visant à développer des stratégies immuno-thérapeutiques pour
-
peptide38
A. M...38
B. Résultats38

2
IV. Bilan du doctorat...39
De 2002 à 2003 : Expérience marquante au cours de ma carrière de chercheur hospitalier :
application potentielle des tétramères CMH-peptide en recherche fondamentale et en
recherche clinique39
De 2003 à maintenant : Résultats marquants obtenus au cours de ma carrière de chercheur à
: 40
I. Problématique de recherche.40
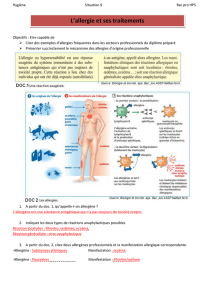
II. Etude des mécanismes des allergies alimentaires...42
A. Action 1 : Etude de la translocation des allergènes à travers l'épithélium
intestinal...43
1. Problématique .43
2. Résultats...44
B. Action 2 natifs ou modifiés sur le système
in vivo ..46
1. Problématique
2.
C. Action 3 : Etude de la phase de déclenchement correspondant à la
dégranulation mastocytaire induite par les allergènes..48
1. Problématique
2. Résultats.....49
D. Action 4 50
1. Problématique 50
2.
E. Conclusion
III. Prévention ou réduction de la réaction allergique par : la stratégie des
53
A. Action 5 : Effet des prébiotiques sur le système immunitaire et la prévention
des allergies.53
1. Problématique 53
2. Résultats......54
B. Projet complémentaire aux prébiotiques: effets des fibres sur le système
immuni.56
1. ...56
2. .....57
C. Conclusion sur la prévention ou la réduction de la réaction allergique par
......58
Projet de recherche original pour l’avenir : « Compréhension des mécanismes
d’interaction de l’aliment (natif, modifié, supplémenté) avec le système immunitaire et la
barrière de l’intestin pour élaborer des stratégies pour agir sur les
allergies. »……………………………………………………………………………………60

3
I. Etude des mécanismes précoces de la réaction allergique engendrés par
60
A. Contexte60
B. Objectifs scientifiques.62
C. Méthodologie
D. Résultats attendus et retombées scientifiques 67
E. Projets envisagés pour réaliser cette étude....68
II. Stratégies nutritionnelles pour réduire ou prévenir les allergies
......................................................................................................69
A. Contexte
B. Objectifs scientifiques...70
C. Méthodologie
D. Résultats attendus et retombées scientifiques .....76
E. Projets envisagés pour réaliser cette étude76
III. Conclusion sur le projet de recherche original..78
Bibliographie………………………………………………………………………………...79
ACTIVITES D’ENCADREMENT…………………………………………………………93
1998 à 2003
2003 à maintenant
Liste des étudiants encadrés.95
I. Post-doctorats ...95
II. Thèses96
III. Masters..96
A. Master
et des Techniques de Nantes.96
B. Master II INAPG- ENSIA Science des Aliments, Massy, ...96
C. Master II BBRT (Biologie, Biotechnologies et Recherche Thérapeutique), UFR de
Nantes96
D. Master II ENSA, Biologie Production animale et Qualité, UFR de Rennes97
E. Master I en « Biologie cellulaire, spécialité physiologie animale », UFR de Rennes..97
IV. Licence professionnelle97
ANNEXES……………………………………………………………………………………99
Annexe 1 : Organigramme de l’unité Biopolymères Interactions et Assemblages…….100
Annexe 2 : Liste des 5 publications représentatives de mes activités…………………...101

4
DOSSIER SCIENTIFIQUE
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
1
/
102
100%