Cours SVT Terminale D: Reproduction, Génétique, Immunité, Évolution
Telechargé par
guy.liamg

Page 1 sur 47 : Cours et exercices de Sciences de la Vie et la Terre – Programme.
1
CLASSE DE TERMINALE D
PARTIE I : UNICITE DES INDIVIDUS ET DIVERSITE GENETIQUE DES POPULATIONS.
Chapitre 1 : Les mécanismes fondamentaux de la reproduction sexuée chez les Mammifères et chez
les Spermaphytes.
Chapitre 2 : Le brassage génétique assuré par la reproduction sexuée et unicité génétique des
individus.
Chapitre 3 : La diversité génétique des populations.
Chapitre 4 : Les prévisions en génétique humaine.
PARTIE II : MECANISME DE LIMMUNITE.
Chapitre 5 : Le soi et le non soi.
Chapitre 6 : Les bases de limmunocompétence Lorigine des cellules immunitaires.
Chapitre 7 : Le déroulement de la réponse immunitaire.
PARTIE III : QUELQUES ASPECTS DU FONCTIONNEMENT DES CENTRES NERVEUX.
Chapitre 8 : Les activités réflexes.
Chapitre 9 : Le fonctionnement des neurones.
Chapitre 10 : Les activités cérébrales et la motricité volontaire.
PARTIE IV : LES REGULATIONS HORMONALES ET NERVEUSES.
Chapitre 11 : La régulation du taux des hormones sexuelles chez lhomme et chez la femme.
Chapitre 12 : La régulation de la glycémie et de la pression artérielle.
PARTIE V : EVOLUTION DE LA TERRE ET DU MONDE VIVANT.
Chapitre 13 : Lhistoire et lévolution de la Terre et des êtres vivants.
Chapitre 14 : Le mécanisme de lévolution.
Chapitre 15 : La lignée humaine.
PROGRAMME DE SCIENCES DE LA VIE ET DE LA TERRE

Page 2 sur 47 : Cours et exercices de Sciences de la Vie et la Terre – Mécanismes fondamentaux de la reproduction
sexuée chez les Mammifères et les Spermaphytes.
2
OBJECTIFS :
A la fin de ce chapitre, lélève sera capable de :
- identifier les organes reproducteurs chez les Mammifères et les Spermaphytes ;
- identifier, expliquer et schématiser les étapes de la méiose ;
- en déduire que le gamète formé est haploïde par rapport à la cellule mère diploïde ;
- dégager les principales étapes de la formation des gamètes ;
- relever la variation du taux dADN par cellule ;
- identifier les différentes phases de la fécondation ;
- en déduire que la fécondation aboutit à la formation dune cellule diploïde appelée zygote à partir de
deux cellules haploïdes ;
- relever les différents mécanismes de la reproduction sexuée que sont la méiose et la fécondation ;
- décrire un cycle de développement à prédominance haploïde, un cycle à prédominance diploïde et un
cycle haplodiplophasique ;
- conclure à travers le cycle chromosomique et le cycle de développement que la méiose et la
fécondation sont deux phénomènes compensateurs qui maintiennent constant le nombre de
chromosomes de chaque espèce.
INTRODUCTION
Chez les animaux, les deux modalités de reproduction (reproduction conforme et reproduction
sexuée) ne coexistent que chez quelques groupes rudimentaires (éponges, hydre, ).
Les êtres vivants les plus évolués (mollusques, insectes, vertébrés, spermaphytes) se
reproduisent exclusivement par reproduction sexuée. Les sexes sont le plus souvent séparés
(gonochorisme ou monoïsme) et on distingue au sein de la même espèce des mâles sécrétant des
gamètes mâles et des femelles sécrétant des gamètes femelles.
Quelques espèces animales (escargots, lombrics) et la plupart des espèces végétales sont
hermaphrodites. Les glandes reproductrices mâles et femelles sobservent chez le même sujet.
1- FORMATION DES GAMETES.
1.1 LES ORGANES REPRODUCTEURS CHEZ LES MAMMIFERES ET LES SPERMAPHYTES.
1.1.1 Chez les Mammifères.
Dune manière générale ; les appareils génitaux sont bâtis suivant le même plan dorganisation chez les
Mammifères quel que soit le sexe. On distingue ainsi :
- les glandes génitales ou gonades ;
- les voies génitales ;
- les glandes annexes.
Les gonades ont une double fonction :
- exocrine par la production des gamètes ;
- endocrine par la production des hormones.
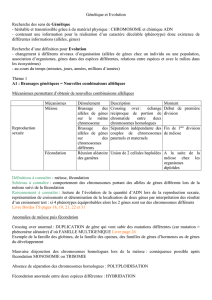
PARTIE I UNICITE DES INDIVIDUS ET DIVERSITE GENETIQUE DES POPULATIONS.
CHAPITRE I LES MECANISMES FONDAMENTAUX DE LA REPRODUCTION SEXUEE CHEZ LES
MAMMIFERES ET LES SPERMAPHYTES.

Page 3 sur 47 : Cours et exercices de Sciences de la Vie et la Terre – Mécanismes fondamentaux de la reproduction
sexuée chez les Mammifères et les Spermaphytes.
3
Figure 1 Figure 2
Annoter les figures 1 et 2 (noter au crayon les gonades, au bic bleu les voies génitales et au bic rouge
les glandes annexes).
Figure 1 : Appareil génital de lhomme. 1- vessie ; 2- Vésicule séminale ; 3- rectum ; 4- canal déférent (spermiducte) ; 5-
épididyme ; 6- prostate ; 7- corps caverneux ; 8- urospermiducte ; 9- corps spongieux ; 10- prépuce ; 11- gland ; 12- orifice
urogénital ; 13- testicule.
Figure 2 : Appareil génital de la femme. 1- oviducte ou trompe de Fallope ; 2- pavillon de loviducte ; 3- ovaire ; 4-
endomètre ; 5- myomètre ; 6- col ; 7- utérus ; 9- petite lèvre ; 10- grande lèvre ; 11- orifice génital ; 12- vulve.
1.1.2 Chez les Spermaphytes.
Lappareil reproducteur des Spermaphytes (Angiospermes et Gymnospermes) est la fleur.
Une fleur complète montre deux types de pièces :
- les pièces stériles et protectrices comprenant le calice (ensemble de sépales) et la corolle
(ensemble de pétales). Les pétales jouent un rôle indirect dans la pollinisation grâce à leurs
colorations vives qui attirent les insectes et les oiseaux. Lensemble des pièces stériles forme le
périanthe.
- Les pièces fertiles comprenant landrocée qui est lensemble des étamines et le gynécée ou
pistil constitué de plusieurs carpelles. Elles produisent les gamètes.
Figure 3
Annoter la figure ci-contre (noter au
crayon les pièces stériles, au bic
bleu les pièces fertiles mâles et au
bic rouge les pièces fertiles
femelles.
Figure 3 : Coupe verticale dune fleur
complète dAngiosperme. 1- grains de
pollen ; 2- anthère ; 3- filet ; 4- étamine ; 5-
stigmate ; 6- style ; 7- ovule ; 8- ovaire ; 9-
pistil ; 10- pétale ; 11- sépale ; 12-
périanthe ; 13- réceptacle ; 14- pédoncule
floral.

Page 4 sur 47 : Cours et exercices de Sciences de la Vie et la Terre – Mécanismes fondamentaux de la reproduction
sexuée chez les Mammifères et les Spermaphytes.
4
1.2- LA MEIOSE : PASSAGE DE LA PHASE DIPLOÏDE A LA PHASE HAPLOÏDE.
La méiose est constituée par une succession de deux divisions cellulaires non séparées par une
synthèse de lADN. Elle réalise la réduction chromatique c'est-à-dire la division par deux du nombre
de chromosomes de la cellule mère. Chaque cellule fille reçoit un seul chromosome de chaque paire de
chromosomes de la cellule mère.
1.2.1 La première division de méiose : la division réductionnelle.
On peut la résumer ainsi quil suit :
On définit les phases suivantes (exemple dune cellule animale à 2n = 4 chromosomes) :
a- Prophase I.
Cest la phase la plus longue de la méiose. On distingue les faits caractéristiques suivants :
1- individualisation des chromosomes à deux chromatides par condensation de la chromatine ;
2- rapprochement puis appariement des chromosomes homologues ;
3- formation des asters à partir du centrosome, début de formation du fuseau de division et
dislocation de lenveloppe nucléaire ;
4- formation des bivalents ou paires de chromosomes homologues appariés ;
5- appariation des figures en X appelées chiasmas au niveau des bivalents.
b- Métaphase I.
Pour chaque bivalent, les centromères des chromosomes homologues se placent de part et dautre du
plan équatorial. Les chiasmas glissent au bout des chromatides : cest la terminalisation des
chiasmas.
c- Anaphase I.
Il y a séparation des chromosomes homologues de chaque bivalent sans clivage ou division des
centromères et migration vers chaque pôle dun lot de n chromosomes à deux chromatides.
d- Télophase I.
Les chromosomes ne se décondensent pas totalement. Il y a séparation des deux cellules filles à n
chromosomes à deux chromatides.
1.2.2 La deuxième division de méiose : la division équationnelle.
On peut la résumer ainsi quil suit :
Les aspects cytologiques de la deuxième division méiotique sont très comparables à ceux dune mitose
ordinaire.

Page 5 sur 47 : Cours et exercices de Sciences de la Vie et la Terre – Mécanismes fondamentaux de la reproduction
sexuée chez les Mammifères et les Spermaphytes.
5
Elle fait immédiatement suite à la première division méiotique et comporte les phases successives
suivantes :
Figure 4 : La première division de méiose : la division réductionnelle.
Justifier le nom de méiose réductionnelle attribué à la première division de méiose au regard de la
figure 4
a- Prophase II.
Elle débute immédiatement après la télophase de la première division méiotique, avec les
chromosomes déjà condensés. Un nouveau fuseau de division se met en place dans chacune des deux
cellules filles. La membrane nucléaire disparaît.
b- Métaphase II.
Les centromères des n chromosomes fissurés se disposent dans le plan équatorial ou plan médian du
fuseau de division.
c- Anaphase II.
Les deux chromatides de chaque chromosome se séparent au niveau du centromère et chacune migre
vers un pôle opposé du fuseau.
d- Télophase II.
Elle conduit à la reconstitution de quatre noyaux fils haploïdes.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
1
/
47
100%