Les enzymes (cours complet)

ENZYMOLOGIE
I) Propriétés générales des enzymes
1.1. Application
- En médecine pour le dépistage des maladies (dosage des prises de sang), par
exemple une augmentation de concentration en amylase dans le sang cause un
disfonctionnement du pancréas
- Outils industriels : lessive, fabrication de papier, de textile, agroalimentaire…
1.2. Propriétés de catalyseur
1.2.1. Efficacité
Un catalyseur accélère une réaction et n’est pas consommé au cours de cette réaction, il est
actif en faible concentration et ne modifie pas le cours thermodynamique de la réaction.
1.2.2. Spécificité
Au niveau des liaisons coupées ou transformées :
- Chimiosélectivité ; ex : protéases qui coupent les liaisons peptidiques NH-CO et
séparent les acides aminés
- Régiosélectivité ; ex : trypsine coupe après un acide aminé basique, chymotrypsine
après un acide aminé aromatique
- Stéréosélectivité ; ex : L-amino-oxydase ou D-amino-oxydase, α ou β
galactosidase, furamase qui réagit sur le furamate en configuration E pour donner
du malate…
1.2.3. Régulation
La concentration en calcium intracellulaire, la phosphorylation ou l’allostérie (modification
de conformation) peut moduler l’activité des enzymes.
1.3. Classification des enzymes
- Les oxydoréductases catalysent le transfert d’électrons et vont avoir comme
substrat l’oxydant et le réducteur. (1)
- Les transférases catalysent le transfert d’un groupe d’atomes R de A vers B ou
inversement. (2)
- Les hydrolases coupent la liaison souvent entre deux carbones ou entre un carbone
et un soufre/oxygène/azote : C—X + H2O → C—OH + HX. Les réactions sont
déplacées vers la rupture de la liaison. (3)
- Les lyases lient deux groupement d’atomes : A=B + X—Y → X—A—B—Y (4)
- Les isomérases permettent la formation d’un isomère d’une molécule donnée :
X—A—B ←→ A—B—X (5)
- Les ligases ou synthétases : produisent l’action inverse des hydrolases. (6)

II) Cinétique des réactions enzymatiques
2.1. Rappel sur la cinétique chimique
La thermodynamique nous renseigne sur l’énergie de l’état initial et de l’état final, ce qui
permet de déterminer si une réaction est faisable, mais ne renseigne pas sur sa vitesse. La
cinétique renseigne sur la vitesse (variation de concentration en fonction du temps). La vitesse
est fonction de la concentration et de la nature du substrat et du produit, de la température, de
la pression, du pH…
!
V=dP
dt ="dS
dt
2.1.1. Cinétique d’ordre 1
Substrat → Produit
!
V=k1S
[ ]
="dS
dt
k1 est la constante de vitesse d’ordre 1 (s-1).
!
ln S
[ ]
S0
[ ]
=k1t
La courbe V = f (S) est une droite de pente k1. T1/2 est indépendant de la concentration
initiale en substrat.
!
T
1/ 2 =ln2
k1
Exemple : saccharose + H2O → glucose + fructose
!
V=k saccharose
[ ]
H2O
[ ]
=k1saccharose
[ ]
2.1.2. Cinétique d’ordre 2
A + B → P
A + B → P + Q
!
V=dP
dt ="dA
dt
!
V=k2A
[ ]
B
[ ]
(k2 en M-1.s-1)
!
1
A0"B0
ln B0
[ ]
A
[ ]
A0
[ ]
B
[ ]
=k2t
si
!
A
[ ]
=B
[ ]
,
!
V=k2A
[ ]
2
!
1
A
[ ]
=k2t+1
A0
[ ]
2.1.3. Cinétique d’ordre 0
V = k0, la vitesse est indépendante de la concentration.
!
V=dP
dt =k0
et
!
P
[ ]
=k0t

2.2. Notion de vitesse initiale
2.2.1. État stationnaire
k1 kcat
E+ S ES E + P
!
V=kcat ES
[ ]
k-1
- Phase pré-stationnaire (ms)
- Phase stationnaire d’ordre 0 (Vi, en s, min) ≠ d’état d’équilibre
- Phase post-stationnaire
Pour une unité d’enzyme : Vi est en µmol/min
Pour un katal (énergie nécessaire pour transformer une mole de substrat, en SI) : Vi est en
mol/s.
2.2.2. Vi = f(E)
Si on fait varier la concentration en enzyme pour une concentration de substrat fixe, la
formation de produit en fonction du temps reste linéaire, plus il y a d’enzyme, plus la réaction
est rapide. Si la concentration en enzyme est exprimée en mg/L, on parle d’activité spécifique
de l’enzyme. La cinétique est d’ordre 1 par rapport à l’enzyme.
!
Vi=k E
[ ]
2.2.3. Vi = f(S)
On fait varier la concentration en substrat pour une concentration en enzyme fixe. La
vitesse initiale augmente de façon parabolique quand la concentration initiale en substrat
augmente.
2.3. Équation de Michaelis Mentes
Elle nécessite plusieurs conditions :
1) la concentration en produit doit être très inférieure à celle en substrat, la
réversibilité de la réaction est ainsi proche de 0
2) la concentration en ES doit être constante au cours du temps
k1 kcat
E+ S ES E + P
k-1
!
Vformation =k1E
[ ]
S
[ ]
!
Vdisparition =k"1ES
[ ]
+kcat ES
[ ]
=(k"1+kcat )ES
[ ]
À l’équilibre :
!
Vf=Vd
donc
!
k1E
[ ]
S
[ ]
=(k"1+kcat )ES
[ ]
!
Et
[ ]
=E
[ ]
+ES
[ ]
donc
!
E
[ ]
=Et
[ ]
"ES
[ ]
!
k1Et
[ ]
"ES
[ ]
[ ]
#S
[ ]
=(k"1+kcat )ES
[ ]

!
V=kcat ES
[ ]
donc
!
V=kcat k1Et
[ ]
S
[ ]
k1S
[ ]
+(k"1+kcat )
!
Vm =kcat Et
[ ]
et
!
Km =k"1+kcat
k1
Donc :
!
V=Vm S
[ ]
S
[ ]
+Km
Cette équation n’est valable que dans les premiers temps de la réaction.
Signification des constantes :
- Km : concentration initiale en substrat pour laquelle
!
Vi=1
2
Vm
- Quand la concentration initiale en substrat est très supérieure à Km,
!
Vi=Vm =kcat Et
[ ]
(kcat est l’efficacité catalytique de l’enzyme, en s-1)
- Quand la concentration initiale en substrat est très inférieure à Km,
!
Vi=kcat
Km
Et
[ ]
S
[ ]
(
!
kcat
Km
est la constante de spécificité, de l’ordre de 108-109)
La comparaison de différentes constantes de spécificité permet de déterminer le meilleur
substrat pour une enzyme donnée.
!
kcat
Km =k1kcat
k"1+kcat
Si kcat est très supérieure à k-1,
!
kcat
Km
tend vers k1.
Représentations linéaires :
- Linewearer-Buck :
!
1
Vi
=f1
S
[ ]
0
"
#
$
$
%
&
'
'
!
1
V=Km
Vm S
[ ]
0
+1
Vm
(voir cours chimie du vivant)
- Eadie-Hofstee :
!
V=fV
S
[ ]
0
"
#
$
$
%
&
'
'
!
Vm =
V S
[ ]
+Km
[ ]
S
[ ]
0
=V+V
S
[ ]
Km
!
V=Vm "Km V
S
[ ]
0

2.4. Des effecteurs de l’activité enzymatique
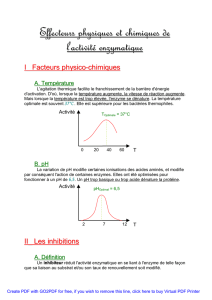
2.4.1. Le pH
- pH optimum entre pK1 et pK2 : forme zwitterion
- dénaturation aux pH extrêmes (réversible ou non), changement de conformation et
perte d’activité
- substrat ionisable : l’activité va dépendre de sa forme ionique (et donc du pH)
Dans la nature, le pH optimum est de 7, celui de la pepsine est de 2 (travail dans
l’estomac).
Pour étudier une activité enzymatique in vitro, on travaille dans un milieu tamponné pour
limiter les fluctuations de pH.
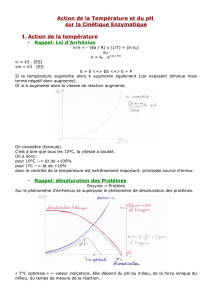
2.4.2. Température
Pour la plupart des enzymes eucaryotes, la température optimale est de 37°C. Il existe
néanmoins des enzymes hyperthermophiles à très haute température optimale pour les
organismes vivants dans des milieux extrêmes.
Activation thermique :
Énergie
état de transition
S ΔG
Enthalpie libre
P
Réaction
!
E
[ ]
=E0
[ ]
e"kt
(kt est le coefficient d’activation thermique)
!
ln E
[ ]
E0
[ ]
=ln V
V0
Energie d’activation : loi d’Eyring
!
k=kBT
h
e"#G$
RT
kB : constante de Boltzmann
h : constante de Planck
 6
7
8
9
10
11
12
6
7
8
9
10
11
12
1
/
12
100%