Corrigé DM1 Concours G2E 2009 Sujet de Biologie 1

1
Corrigé DM1 Concours G2E 2009
Sujet de Biologie 1
QUELQUES ASPECTS DU DEVELOPPEMENT PRECOCE DE LA PLANTE
CHEZ LES ANGIOSPERMES
Les passages en italiques sont extraits du rapport du jury, téléchargeable sur le site du concours.
Introduction et conclusion étaient exigés par le sujet. En 2008, au contraire, le sujet précisait
qu’elles n’étaient pas nécessaires. Il faut donc s’adapter et faire ce qui est demandé.
Introduction.
- Définir « développement », « Angiospermes ».
- Quelques questions posées par le sujet : que deviennent les zygotes formés lors de la double
fécondation ? Comment se forme la graine ? Quels sont les liens trophiques entre la graine en
cours de formation et la plante mère ?
- Présentation du plan : énumération des 3 axes abordés dans les 3 parties.
1. Le développement de l’embryon.
1.1 Événements cytologiques de la double fécondation : voir cours B5
Conséquences génétiques : formation d’un zygote diploïde (qui donnera l’embryon) et d’un zygote
triploïde (qui formera l’albumen).
Dans le cas du croisement cité dans l’énoncé, en supposant que la présence du pigment dans la
couche à aleurone est spécifiée par 1 gène à 2 allèles, notés b (pas de pigment) et B (présence
d’un pigment), les pieds descendants des 2 variétés de maïs forment des grains dont l’albumen
triploïde a pour génotype Bbb. Celui-ci synthétise le pigment dans sa couche à aleurone, ce qui
montre que l’allèle B est dominant.
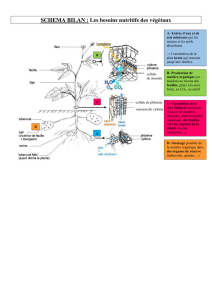
1.2 Document 1a (à gauche) : embryon globuleux ; embryogenèse précoce.
1 : albumen. 2 : embryon. 6 : suspenseur. 7 : nucelle.
Document 1b (à droite) : embryon cordiforme ; organogenèse.
1 : albumen 3 : méristème apical racinaire (MAR). 4 : méristème apical caulinaire (MAC).
5 : 2 cotylédons. Devenir des structures
Après la fécondation Après développement
albumen Accumulation de réserves utilisées lors du DE
(graine exalbuminée) ou à la germination
embryon plantule
MAR radicule
MAC tigelle + gemmule
cotylédons Accumulation de réserves utilisées lors de la
germination si graine exalbuminée ou feuilles
cotylédonaires si graine albuminée
suspenseur disparaît
nucelle disparaît
1.3 2a : albumen ; 2b : couche à aleurone. Ces structures dérivent de l’œuf triploïde.
2c : enveloppe du caryopse = péricarpe (dérive de la paroi ovarienne) et tégument de la graine
(dérive des téguments ovulaires).
2. Le développement de la graine
2.1 Localisation des réserves
A l’échelle de la semence A l’échelle cellulaire
Céréales
(blé, seigle, orge, maïs) albumen
Légumineuses
(pois, fève, haricot, lupin) cotylédons amyloplastes (amidon) ;
grains d’aleurone (protides) ;
noix cotylédons
ricin albumen gouttelettes cytosoliques
(lipides)

2
2.2 a) Le document 4 montre que le poids de l’embryon (exprimé en % d’une valeur maximale ?)
croît suivant une fonction affine du temps, de 30 à 100 JAP, c'est-à-dire en même temps que se
fait l’accumulation des réserves dans l’albumen.
Deux origines peuvent être envisagées : une synthèse propre à la graine, ou l’importation de
réserves synthétisées par la plante-mère.
Dans les 2 cas, c’est la photosynthèse de la plante-mère qui produit la matière organique.
Les protéines de réserve sont synthétisées à la suite de l’expression des gènes qui les codent. Les
triglycérides (lipides de réserve) proviennent de l’estérification du glycérol par 3 acides gras. Les
acides gras sont eux-mêmes synthétisés à partir de l‘acétyl-coenzyme A.
Dans tous les cas, la mise en réserve est la conséquence de l’expression d’un programme
génétique particulier.
b) Sur le document 4, la phase de déshydratation commence après la fin de la mise en réserve. En
effet, lorsqu’elle est achevée, elle entraîne l’arrêt de l’activité métabolique (vie latente). Les
membranes sont protégées des effets de la déshydratation par la synthèse de protéines stabilisant
leur structure.
La déshydratation est un facteur de protection contre le gel. Plus une semence est déshydratée,
plus sa longévité est grande.
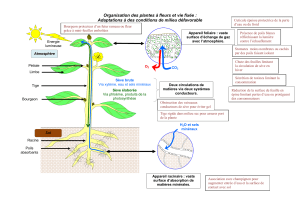
2.3 Un organe source est à l’origine du flux de matière organique de la sève élaborée. Il s’agit le
plus souvent des feuilles, organes photosynthétiques, mais lors de l’hydrolyse de ses réserves, un
organe de réserve (tubercule de pomme de terre en germination, par exemple) joue le rôle d’un
organe source.
Un organe puit est destinataire du flux de la sève élaborée : il peut s’agir d’organes de réserve,
d’organes en croissance, des fleurs, des fruits.
2.4 L’insolubilité de l’amidon pose le problème de sa mobilisation et de son transport. L’amidon
synthétisé dans le chloroplaste est hydrolysé avant transport vers les organes puits. La sève
élaborée contient des polyosides solubles (principalement le saccharose, diholoside)
3. Le transport de la sève élaborée
3.1a) Annotation du document 5a.
A : cellule compagne. B : paroi de la cellule criblée. C : cytoplasme de la cellule criblée. D : crible
formé de pores.
Les tubes criblés conduisent la sève élaborée des organes sources aux organes puits. Les cellules
compagnes ont un double rôle : par leurs divisions, elles peuvent former de nouveau tubes
criblés ; ce sont des cellules de transfert, à l’origine de la charge active des molécules organiques
dans le phloème.
b) « L’organisation des tubes criblés du phloème et leur différenciation ont été traités par analogie
avec les tubes du xylème. L’erreur la plus fréquente restant que l’on nous présente des cellules
mortes. »
C’est logique car la différenciation des tubes criblés n’est pas au programme
c) Voir schéma complété page 3.
« Le schéma à compléter n’a pas apporté son lot de satisfaction car pour de nombreux candidats
la voie apoplastique passe en dehors de la paroi cellulaire, les flèches sont inversées (le flux de
sève allant des tubes criblés aux cellules du mésophylle), et la charge de la voie apoplastique à
partir de la voie symplastique est omise. »
3.2
- expérience 1 : tant que les cellules plongées dans le mannitol restent turgescentes, cela signifie
que leur potentiel osmotique est plus faible que celui de la solution de montage. Les cellules du
phloème ont donc un potentiel osmotique plus faible que les autres cellules de la feuille, sans
doute à cause d’une concentration élevée en saccharose. Cela semble montrer qu’il existe une
membrane entre le complexe phloémien et les autres tissus de la feuille (ce que ne montre pas le
document 5).
- expérience 2 : l’apparition du saccharose dans la solution indique qu’il a été libéré dans
l’apoplasme. Ceci confirme l’hypothèse précédente. La charge du phloème est précédée d’un
passage des molécules organiques dans l’apoplasme.
3.3 Les observations rapportées montrent :
- que l’absorption du glucose est un processus actif spécifique (n°4 et 5) ;
- qu’il s’agit d’un symport saccharose / protons (n°3)

3
- que le cation transporté avec le saccharose est H
+
(n°1 et 2)
Ceci conduit à construire un schéma représentant l’existence de deux transports actifs (transport
actif primaire par la pompe à protons, transport actif secondaire de type symport saccharose/H
+
).
3.4 L’observation des stylets de pucerons montre que la sève élaborée est sous pression dans les
tubes criblés du phloème. Les valeurs du tableau permettent de calculer une différence de
pression de turgescence de 4 bars entre les organes sources et les organes puits. C’est cette
différence de pression qui est le moteur principal de circulation de la sève élaborée.
Elle s’explique par l’absorption active du saccharose par les cellules du phloème qui en abaisse le
potentiel osmotique, engendrant une absorption osmotique d’eau à partir du xylème qui augmente
la pression de turgescence. Le phénomène inverse s’observe au niveau des organes puits : la
décharge du phloème entraîne l’eau hors des tubes criblés.
Un autre mécanisme se rajoute à ce déplacement en masse de la sève élaborée. La
concentration de saccharose de la sève est plus élevée au niveau des organes sources que des
organes puits. Cette différence permet une diffusion (lente, comme toute diffusion) du saccharose
des organes sources aux organes puits.
3.5 Les faits rapportés suggèrent l’existence de trois mécanismes de décharge du saccharose.
- Un premier mécanisme de décharge indépendant de transporteurs de saccharose et de source
énergétique (fait n°2), donc lié à un gradient de c oncentration, passe par la voie symplastique
(fait n°1).
- Un deuxième est lié à une hydrolyse du saccharose en glucose et fructose par une invertase
acide (fait n°3) suivie d’un transport spécifique du glucose et du fructose (fait n°4).
- Un troisième dépend d’un processus consommateur d’oxygène et d’une ATPase et de
transporteurs actifs spécifiques du saccharose ; il est donc lié à une dépense énergétique.
Ces deux derniers mécanismes passeraient par voie apoplastique.
Document 5b : du mésophylle à la cellule criblée.
. D’après Campbell – Biologie – De Boeck Université – 1995.
Conclusion
- Résumé des points abordés.
- Ouverture scientifique : les dormances, caractéristiques des semences des Angiospermes.
- Ouverture pratique : importance des semences pour l’alimentation, pour l’agriculture ; aspects
économiques de l’approvisionnement en semences des agriculteurs, notamment dans les pays
émergents.
voie symplastique
passage par la voie apoplastique (= «charge de la voie apoplastique à partir de la
voie symplastique » ?)
transport actif
1
/
3
100%