– UEI : Sciences biologiques, pathologiques, sémiologie–

2015-2016 Métabolisme des hormones Stéroïdes (2)
Semaine :
Date :
Heure : de
Professeur :
Binôme : Correcteur :
Remarques du professeur:
!"
# $%
# & ' % $$(%
!)"!
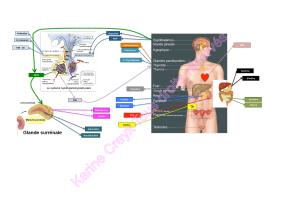
A) Site de synthèse de l'aldostérone
B) Biosynthèse de l'aldostérones
C) Sécrétion et transport plasmatique de l'aldostérone
D) Fonctions biologiques et mode d'action de l'aldostérone
E) Régulation de la sécrétion de l'aldostérone
F) Catabolisme de l'aldostérone
# &'%$(% !")!
A) Site de synthèse du cortisol
B) Synthèse du cortisol
C) Sécrétion et transport du cortisol
*+,

2015-2016 Métabolisme des hormones Stéroïdes (2)
# & ' % $$(%
!)"!
A) Site de synthèse de l'aldostérone
! "#$ %%&!! ! !"$''' !(
)!*!!'+ ! ,-+
+ ! ,!!#.!&!*!!'/00
0
1 '!+(+-2
–3+2
–3!(2 ! ,
4 !(!+-'% 562
–7+
–7%
–7!
3 -! ' ( '! %%!'*!8!"
! "!'! &!!*!!'-$
B) Biosynthèse de l'aldostérone
1) 1ère étape de Biosynthèse
3!!!'%$.9'! !#!' 6 ':
* # !$ ! ;!'! !!' *!'
$;! # !''!;!+< !'!
% !''!".
; ;!"!=#+81!"
!' $ ;! ;,>6&$!$
!#:###
4! #!*' $+!';2
/0%$10
9 '!!8 ; # $+ ?!
$ $#%!!; ! ;% !9@#' ( ;
!''*+'!9@%()%! ;' !%!*(*'
!*(*''*!%!+< &AB'!!+'!''!!8
!+!%#%(#6$!!! ! ;!&! ;!
2+,

2015-2016 Métabolisme des hormones Stéroïdes (2)
( ;! !!' ' $!
!10%
C%!$-*6 $! ;3! &! 23422
%5%!'! '(3DA
! %!!$;(!!!#'!! !
! ! 6 !6 !!$;
'+ !!$8;! !''!"1!+ !
+!'!'!'+ ! ,!+
EFGHIJKLMGHNGHOJHKPQROJLMSFHNGHOJHTMSUVFLWXUGHNGHOYJONSULPKSFGHIJUUGHIJKHRFGHPLJIGHNGHKPQROJLMSFHNGHZL[\]H^YGUL_`_H
NMKGHRFGHPLJIGHNGHKPQROJLMSFHNRHLKJFUaGKLHNRH^WSOGULPKSOHNRH^VLSIOJUbGHcGKUHOJHbJLKM^GHbMLS^WSFNKMJOGdH
3 $+!!! $+(*!#. '+& !&(*!
;)! !%&% ! !
'!%6'!: * %#&''$.
2) 2ème étape de la biosynthèse
9(*!% !9@ '!'3#% ! !
6 ''!9' !
;'! * !!3!3
;'!!' * A#
! ;:!'e!'!!:
&; !! !&(*!!'644
0%$104$(%4%0%$.3!!-**+8
'*(*3
'!%!$$.
3!!-*!%'!'+#-
+ ! C&! $!$
$$%7$%$5'
'$!
3&$ $ '+! !'%
$
3) 3ème et 4ème étapes de la biosynthèse
C!'6 !&*(*!3'+!'
&-*2*40%$10
'%&''*(*'+!>**4
%10$$ ! !&*(*!
3'**440%$10'%$$
' % !9@ ;!
4) 5ème étape de la biosynthèse
3!!!''!% !&!! 6.!
9*(*!3=#'*840%$10! %!&% ! 4
&!!$% !*3=#'%%!*(*3=
A'8*(*!3=#(*!% ! '*8410%3!!8!
!!*+% !*# !!6&!'*840%$104
$$
!$!-*!6=,*(*!=,(*'! '!%! '(
6+,

2015-2016 Métabolisme des hormones Stéroïdes (2)
&!*!C!' %6-*6&!$-$'
C) Sécrétion et transport plasmatique de l’aldostérone
•1 !6!2=,5
•3 !!'!6*'2f:'
!'!&%2
–g%;# !$
–g%%;!&;# !$
!'!'!6' %6&! '!!+# 6 !
6%; !$!!8;!
D) Fonctions biologiques et mode d'action de l'aldostérone
B! &+'8!' '! .!#!!!'
;
&!!$ ;+6 !; !!'
% !&!!'!!'+#&;'!!;'!h0 !
8!;56& '!!(! '!6+
i.!0$9# !0$$#+! !!
&+
C;;!&'!;;.!e!#$!
$ !+
i&;;&!'# &!%$
i&!0$02+!!$+ ! 6&''
*'$
C;!'!&' !@+
&$! '!!&!$
!656*'%(!
'+8 ; ! (!
'!#'!!!.!!!
'!
1&! j! ! !6!!!!$
'''!6%!!'!
6#hkAB
i&!0:
E) Régulation de l'aldostérone
1.!*'!; !!.!!.*'!5
!&!!+!
'!&!!
'!*!8$
%$# !;!+!
'!
3!!' '!$!'!!
;+,

2015-2016 Métabolisme des hormones Stéroïdes (2)
'l'; '!$ 5(!,+
!'!! !'3!!-* $'!!''!
WPIJLMmRGHnH<=>?@ABCD?EA?B@F?DG'%''!' !# !%27*]H^OMcPGHIJKH7-0
%9$'72''! ' !# !$
& !'!*'!#(!*!;'2
–4 9$2!8!!8
o623&!%%!6!&+!))C $ ! !'
+&*'!'$ ! !
–$9# &!+!$ !
o62pHOSFQHLGKbG]HISRKHIKSOSFQGKH^GLLGHJ^LMSFHURKHOGUHcJMUUGJRq]HOrJFQMSLGFUMFGHsHNP^OGF^WGHOJH
UVFLWXUGHGLHIGKbGLHOJHUP^KPLMSFHNYJONSULPKSFGHmRMHJRQbGFLGHOJHcSOPbMGdH
F) Catabolisme de l'aldostérone
36!'!'&!!$;'!"
3 !;!'70#'!'!!&!
!; !'! '8!!& ''!% !!
.!&&!!!!!
!! !!&!*';;! !;!! &
!8*'8%
o'*' !!2
*# o; * !&*'
2# o !% ! !
6# 1% ! %(! *';!&!
!8*' & + 6%!+ 5+
•B!.; !% ! !
•35+& + 66$%(
9@
•C*;;6
!.*'2&!!$ % !
!#% !9@!;
4<=HIJ>CBKLCDMG^YGULHJMFUMHOJHIKGbMXKGHKPJ^LMSFHNR
^JLJTSOMUbGHnHSFHKPNRMLHOJHNSRTOGHOMJMUSFHGFHISUMLSFHtu
* 'NOPOBQLRBSIRTKC>ED
•v(!*+86 ;!'
%%0%$%$
•)!% ! !'!!+
% !9@ !!'!
◦9!% ! !3% !9@$;*!8#'64U40%$104
ECISBVRDGBQLRBSIRTKC>EDG'%CICS>OGHLRSBO><RBECISB?D
◦9!% ! !3% ! 'WXOHLRSBQLOECISBVRDGBQLRBSIRTKC>ED
wGH^SbISUPHaSKbPHGULHOGHHDQ>HLRSBQL><RBECISB?D
Y+,
 6
7
6
7
1
/
7
100%