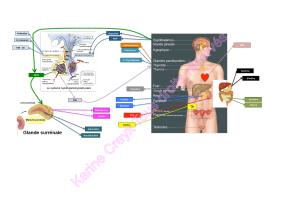
Télécharger le fichier - Fichier

2015-2016 Métabolisme des hormones Stéroïdes (2)
Semaine :
Date :
Heure : de
Professeur :
Binôme : Correcteur :
Remarques du professeur:
!"
# $%
# &'%$$(%
!)"!
#%*+%,%$
#*+%,%$
#$$$%,%$
#-%%,%,%$
#"%$%,%$
-# %,%$
#&'%$(% !")!
#%*+%$
#*+%$
#$$$%$
./0

2015-2016 Métabolisme des hormones Stéroïdes (2)
#&'%$$(%
!)"!
# %*+%,%$
! "#$ %%&!! ! !"$'''
!()!*!!'+ ! ,-+
+ ! ,!!#.!&!*!!'
/000
1 '!+(+-2
–3+2
–3!(2 ! ,
4 !(!+-'% 562
–7+
–7%
–7!
3 -! ' ( '! %%!'*!8
!"
! "!'! &!!*!!'1$
# *+%,%$
.2.+$%*+
3!!!'%$#9'! !#!' 6 '
: * # !$ ! ;!'! !!'
*!'$;! # !''!;!+<
!'!% !''!"#
; ;!"!=#+81!"
!' $ ;! ;,>6&$
!$ !#:###
4! #!*' $+!';2
3*%$4*
9 '!!8 ; # $+ ?
!
$ $#%!!; ! ;% !9@#' (
;!''*+'!9@%()%! ;' !%!
*(*'!*(*''*!%!+< &AB'!!
+'!''!!8 !+!%#%(#6$!!! ! ;!
&! ;!( ;! !!' '
$!
5/0

2015-2016 Métabolisme des hormones Stéroïdes (2)
!4*%
C%!$-*6 $! ;3! &! 567
55%8%!'! '(3DA
! %!!$;(!!!#'!!
!! ! 6 !6 !!$;
'+ !!$8;! !''!"1!+
!+!'!'!'+ ! ,!+
EFGHIJKLMGHNGHOJHKPQROJLMSFHNGHOJHTMSUVFLWXUGHNGHOYJONSULPKSFGHIJUUGHIJKHRFGHPLJIGHNGHKPQROJLMSFHNGHZL[\]H
^YGUL_`_HNMKGHRFGHPLJIGHNGHKPQROJLMSFHNRHLKJFUaGKLHNRH^WSOGULPKSOHNRH^VLSIOJUbGHcGKUHOJHbJLKM^GH
!
3 $+!!! $+(*!#. '+& !&(*!
;)! !%&% ! !
'!%6'!: * %#&''$#
525+%*+
9(*!% !9@ '!'3#% !
! 6 ''!9
' !;'! *
!!3!3;'!!
' * A#! ;:!'d!
'!!:
&; !! !&(*!!'977
*%$4*7$(%7%*%$#3!!-**+8
'*(*3
'!%!$$#
3!!-*!%'!'+#
-+ ! C&!
$!$$$%,$%$
8' '$!
Cela n'à rien à voir ici avec la progestérone sécrétée par le
follicule ovarien.
929+:+%*+
C!'6 !&*(*!3'+!
'&-*5.7*%$4*
'%&''*(*'+!>..7
%4*$$ ! !&*(*!
3'..77*%$4*'%$$
' % !9@ ;!
:2;+%*+
3!!!''!% !&!! 6
.!
9*(*!3=#'.<7*%$4*! %!&% ! 4
&!!$% !*3=#'%%!*(*3=
A'8*(*!3=#(*!% ! '.<74*%3!!8!
!!*+% !*# !!6&!'.<7
*%$4*7$$
9/0

2015-2016 Métabolisme des hormones Stéroïdes (2)
!$!-*!6=,*(*!=,(*'! '!%! '(
&!*!C!' %6-*6&!$-$
'
# $$$%=%$
•1 !6!2=,5
•3 !!'!6*'2e:'
!'!&%2
–f%;# !$
–f%%;!&;# !$
!'!'!6' %6&! '!!+#
6 !6%; !$!!8;!
# -%%,%,%$
B! &+'8!' '! .!#!!!
';
&!!$ ;+6 !; !!'
% !&!!'!!'+#&;'!!;'!g0
!8!;56& '!!(! '!6+
h.!*$># !*$$#+! !!
&+
C;;!&'!;;.!d!#
$!$ !+
h&;;&!'# &!%$
h&!*$*2+!!$+ ! 6&
''*'$
C;!'!&' !
@+
&$! '!!&!
$!656*'%(
!'+8 ; !
(!'!#'!!!.!
!!'!
1&! i! ! !6!!!
!$'''!6%!!
'!6#gjAB
h&!*?
# "%,%$
1.!*'!; !!.!!.*'!5
!&!!+!'!&!!'!
*!8$%$# !;!+!'!
:/0

2015-2016 Métabolisme des hormones Stéroïdes (2)
3!!' '!$!'!
!'k'; '!$
5(!,+
!'!!
!'3!!-* $
IGLMLHIGILMNGHWPIJLMlRGHmH@ABCDEFGHCIECFDJCHK'
%''!' !# !%2,
.]H^OMcPGHIJKH,1*%>$'
,5''! ' !# !$
& !'!*'!#(!*!;'2
–4 >$2!8!!8
n623&!%%!6!&+!))C $ ! !
' +&*'!'$ ! !
–$># &!+!$ !
n62oHOSFQHLGKbG]HISRKHIKSOSFQGKH^GLLGHJ^LMSFHURKHOGUHcJMUUGJRp]HOqJFQMSLGFUMFGHrH
NP^OGF^WGHOJHUVFLWXUGHGLHIGKbGLHOJHUP^KPLMSFHNYJONSULPKSFGHlRMHJRQbGFLGHOJHcSOPbMGsH
-# %,%$
36!'!'&!!$;'!"
3 !;!',*#'!'!!&!
!; !'! '8!!& ''!
% !!.!&&!!!!
!!! !!&!*';;! !;
!! & !8*'8%
n'*' !!2
.2 n; * !&*'
52 n !% ! !
92 1% ! %(! *';!&
! !8*' & + 6%!
+ 5+
•B!.; !% ! !
•35+& + 66$%(
9@
•C*;;6
!.*'2&!!$
% ! !#% !9@!;
4@ALMNBGFOPGHQK &!
OJHIKGbMXKGHKPJ^LMSFHNRH^JLJTSOMUbGHmHSFHKPNRMLHOJ
;'!t: * 'R7:7
FSPTFUMTVOGBIH
•u(!*+86 ;!
;/0
 6
7
6
7
1
/
7
100%