04 02 2016 9h00 10h00 paumelle

2015-2016 Prolifération et apoptose
Contrôle du cycle cellulaire
Biologie cellulaire
– UE 7: – Sciences biologiques
Diapos à venir sur Moodle
Semaine : n°2 (du 01/02/2016 au
05/02/2016)
Date : 04/02/2016
Heure : de 9h00 à
10h00 Professeur : Pr. Paumelle
Binôme : n°37 Correcteur : n°39
Remarques du professeur
•Le professeur s'étonne de ne pas avoir beaucoup d'étudiants en cours
•Elle hésite à mettre les diapos sur Moodle
•L'année dernière elle a refusé de donner une correction en amphi d'un sujet d'annales car il n'y avait
pas assez d'étudiants en cours
•Elle ne sait pas si elle va le faire cette année, cela dépend du nombre d'étudiants qui viennent en cours
prochainement
PLAN DU COURS
I. Destin cellulaire physiologique et pathologique
II. Les protéines de régulation du cycle cellulaire
A. Les complexes cyclines/CDK
1. Définition
2. Rôle
B. Régulation des cyclines
1. Régulation de leur synthèse
Régulation transcriptionnelle
Rôle des facteurs de croissance / mitogènes
2. Régulation de leur dégradation
Phosphorylation, Ubiquitinylation, Protéasome
3. Régulation de leur localisation cellulaire
Cycline E et A nucléaires
Cycline B cytoplasmique ou nucléaire
1/16

2015-2016 Prolifération et apoptose
Contrôle du cycle cellulaire
I. Destin cellulaire physiologique et pathologique
Le devenir de toute cellule se résume à 3
possibilités :
–Prolifération/croissance
–Différenciation (adopter des fonctions
spécialisées)
–Apoptose
Contrairement aux bactéries qui se multiplient seulement quand il y a des nutriments, les cellules de
l'organisme vont se multiplier et se diviser seulement s'il y a besoin d'un apport de cellules dans
l'organisme.
Les facteurs de croissance et les facteurs mitogènes vont permettre d'induire la division cellulaire si
nécessaire.
Ces 3 processus sont très contrôlés, que ce soit lors de l'embryogenèse ou chez l'Homme adulte.
Il y a une homéostasie entre ces différents processus jusque l'âge adulte.
Chez l'adulte on observe moins de divisions cellulaires car les membres sont formés donc on n'a pas
besoin de division cellulaire. On fabrique encore chez l'adulte plusieurs milliers de cellules, comme les
cellules sanguines ou les cellules de l'épiderme par exemple qui doivent être renouvelées. Certaines
cellules sont renouvelées dans l'organisme mais pas toutes (les cellules cardiaques et les cellules
nerveuses ne se renouvellent pas).
Il faut qu'il y ait un contrôle entre ces processus biologiques. Si ces contrôles sont perturbés, cela peut
conduire à des pathologies comme des cancers, qui se développent suite à un défaut ou une
augmentation de prolifération, d'apoptose ou de différenciation.
Si l'on a un tissu qui se divise normalement et qui meurt normalement (quand c'est nécessaire) à ce
moment là l'homéostasie est contrôlée et on a une croissance de l'organisme qui est normale.
Si on a par contre une perturbation au niveau de la division des cellules suite à la mutation des gènes, et
que les cellules se divisent constamment, même avec une apoptose normale, cela peut conduire à la
prolifération d'un cancer.
2/16

2015-2016 Prolifération et apoptose
Contrôle du cycle cellulaire
Si on a une croissance et une division des cellules qui sont normalement contrôlées mais que l'on a un
processus d'apoptose inactif, il y a un risque que ces cellules deviennent tumorales et cancéreuses.
→ Une cellule devient cancéreuse par altération d'au moins un de ces 3 processus.
Ces 3 processus sont contrôlées par le cycle cellulaire. Le cycle cellulaire des cellules eucaryotes a 4
phases (G1, S, G2, M) et il y a des points de contrôles au niveau de ces phases : en phase G1, en phase
G2 et en phase M.
Les points de contrôle sont des endroits où la cellule vérifie son environnement et vérifie si tout son
ADN est répliqué pour continuer de progresser dans le cycle cellulaire et de produire des cellules
identiques à la cellule mère pour conserver l'information génétique.
La cellule peut décider de s’arrêter dans son cycle pour essayer de réparer l'ADN si nécessaire, ou se
différencier, surtout en phase G1 où la cellule vérifie l'environnement et la qualité de son gène.
Si on n'a pas assez de facteurs mitogènes, la cellule rentre en phase G0 ou phase de quiescence, et
attend de s'orienter vers un autre programme, rentre en apoptose, ou encore attend que d'autres facteurs
mitogènes soient produits.
En phase G2, une fois que la réplication est terminée, il y a vérification de l'intégrité de l'ADN, s'il y a
des erreurs, la cellule les répare ou s'il y a impossibilité de réparation, les cellules vont mourir par
apoptose.
Le point de contrôle en phase M est le moment où la cellule vérifie que tous les chromosomes sont
associés aux microtubules pour permettre la bonne séparation des chromosomes. A ce moment là les
cellules peuvent aussi mourir par apoptose.
Des signaux intracellulaires peuvent influencer la réponse de la cellule.
II. Les protéines de régulation du cycle cellulaire
On a identifié les cyclines et les CDK en utilisant génétiquement l'oeuf de Xénope et la levure.
On a essayé ensuite de comprendre comment elles étaient capables de contrôler le cycle cellulaire.
A. Les complexes cyclines/CDK
1. Définition
KINASES-CDK
Les kinases CDK (Cyclin dependant kinases) sont au nombre de 13 chez l'Homme. Ce sont des sérine/
thréonine kinases. Ce sont des protéines qui vont phosphoryler sur des résidus sérine et thréonine.
Elles ont une activité dépendante de l'association aux cyclines via leur extrémité Nter.
L'activité des CDK est également dépendante de la phosphorylation. Elles-mêmes sont phosphorylées
sur des résidus tyrosines et thréonines.
Elles phosphorylent et régulent l'activité de nombreuses protéines impliquées dans la progression du
cycle cellulaire.
3/16

2015-2016 Prolifération et apoptose
Contrôle du cycle cellulaire
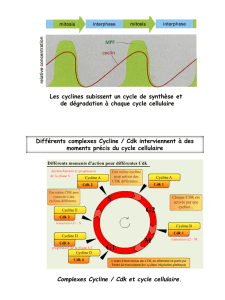
CYCLINES
Les cyclines sont au nombre de 25 mais ne sont pas toutes impliquées dans le cycle cellulaire.
Celles impliquées dans le cycle cellulaire possèdent toutes une région commune d'interaction avec les
CDK que l'on appelle la cycline box. Ce sont des protéines régulatrices des CDK. Quand elles sont
exprimées à un taux important elles permettent de former le complexe actif cycline/CDK.
L'expression de ces protéines est cyclique : diminution ou augmentation de leur expression selon la
phase du cycle cellulaires.
Au niveau des différentes phases du cycle cellulaire, différents complexes cyclines/CDK se forment et
permettent aux cellules de progresser dans le cycle :
–Cycline de G1 : Cycline D couplée avec CDK4 ou CDK6
–Cycline de la transition G1/S : Cycline E couplée avec CDK2
–Cycline de S : Cycline A couplée à CDK2
–Cyclines mitotiques : Cycline A puis B couplées avec CDK1
Ces différentes cyclines donnent une spécificité d'action.
On a également une cycline régulatrice : Cycline H couplée à CDK7.
Cette cycline est importante car elle est responsable de la phosphorylation des CDK.
Pour que les CDK soient actives, il ne faut pas seulement qu'elle se lient aux cyclines mais il faut
également qu'elles soient phosphorylées.
Le complexe cycline H/CDK7 est tout le temps formé dans la cellule. Il ne subit pas de régulation
cyclique.
Il a une activité constante contrairement aux autres cyclines qui agissent seulement transitoirement dans
une phase du cycle et deviennent inactives après. Le complexe Cycline H/CDK7 est toujours actif.
Résumé de l'intervention des complexes cyclines/CDK lors du cycle cellulaire :
La fluctuation de l'activité des complexes cyclines/CDK est responsable de la progression du cycle
cellulaire.
4/16

2015-2016 Prolifération et apoptose
Contrôle du cycle cellulaire
2. Rôle
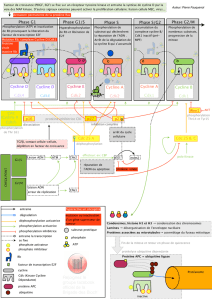
Les cyclines de G1 ou G1/S jouent un rôle important dans le passage du point de restriction, qui est le
point de contrôle en G1 et qui permet à la cellule de pouvoir effectuer un cycle cellulaire complet.
Il faut que les cellules passent ce point de contrôle pour progresser dans le cycle cellulaire.
Pour passer ce point de contrôle, la formation du complexe cycline D/CDK4 (qui est une cycline de G1)
va aller phosphoryler la protéine du rétinoblastome (qui est un inhibiteur du cycle cellulaire, et qui va
bloquer le cycle cellulaire) car cette protéine séquestre un facteur de transcription E2F.
Lorsque le complexe cycline D/ CDK4 se forme, on a phosphorylation de la protéine du rétinoblastome
par CDK4 ou CDK6. Cette phosphorylation va permettre à E2F d'être libéré et de se fixer sur l'ADN
pour induire la transcription des gènes et notamment celle de PCNA (activateur de l'ADN polymérase
delta). E2F peut également induire l'expression d'autres cyclines comme la cycline E ou la cycline A.
La cycline E va se coupler à la CDK2 et va être également capable de phosphoryler la protéine du
rétinoblastome pour avoir le temps d'induire la transcription de gène qui permet à la cellule de passer en
phase S.
Une fois la cellule en phase S, E2F induit
l'expression de la cycline A qui est la
cycline de la phase S.
La cycline A se couple à la CDK2 et
favorise au niveau de l'ADN le recrutement
de PCNA qui permet d'induire la réplication
de l 'AD N pa r l 'a ct iv at io n d 'A DN
polymérase.
5/16
 6
7
8
9
10
11
12
13
14
15
16
6
7
8
9
10
11
12
13
14
15
16
1
/
16
100%