fig cours 4 - WordPress.com

Les cyclines subissent un cycle de synthèse et
de dégradation à chaque cycle cellulaire
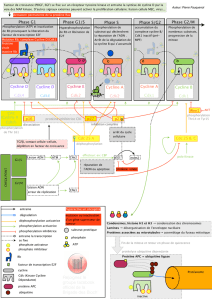
Différents complexes Cycline / Cdk interviennent à des
moments précis du cycle cellulaire
Complexes Cycline / Cdk et cycle cellulaire.

DDCP = DNA
Dammage Checkpoint
RCP = Replication Checkpoint
MPC = mitotic Checkpoint

La voie Ras - MAPKinase permet,
suite à la stimulation par des
facteurs de croissance, de
déclencher l'expression du gène
codant la Cycline D.
Les traductions de Myc et de la
Cycline D ont lieu dans le cytoplasme.
La transition de l’état de repos vers la prolifération
Le point de restriction, limite entre G1 précoce et G1 tardif

Le passage de G1 à S
Action Cycline D / Cdk4 sur la protéine Rb
(rétinoblastome)
Une fois activé, le complexe Cycline D / Cdk4 peut
phosphoryler la protéine Rb.Ceci permet alors la
libération par cette dernière du facteur de transcription
E2F. Une fois libéré, E2F se fixe sur l'ADN, activant
l'expression de gènes codant pour des protéines
nécessaires au déroulement de la phase S.

Une boucle de rétro-activation prépare le passage du
point de restriction
)
La phase G1 prépare la phase S. L'activation du facteur de
transcription E2F, grâce à la phosphorylation de la protéine Rb, permet
la synthèse de gènes de la phase S. La transcription de Cdk 2 et de
Cycline E, en particulier, permet d'augmenter cette activation de E2F,
ce complexe phosphorylant Rb .Ce mécanisme permet à la cellule de
préparer son entrée en phase S.
 6
7
8
9
10
11
12
13
6
7
8
9
10
11
12
13
1
/
13
100%






![(Microsoft PowerPoint - LSV1 CM XI 16 net [Mode de compatibilit\351])](http://s1.studylibfr.com/store/data/001219303_1-c93358a33819374b37626757f18c51e2-300x300.png)



