ovv p2 c2 developpement vegetatif compressed

ORGANISATION DU VIVANT VÉGÉTAL!
PARTIE 2: REPRODUCTION VÉGÉTALE - DE LA FLEUR À LA GRAINE (EB)!
!CHAPITRE 2: DÉVELOPPEMENT
VÉGÉTATIF!
!
I - APPAREIL VÉGÉTATIF
!A) Introduction aux Angiospermes: phylogénie
•Dans les Spermatophytes (plantes à graines = plantes à spores), on s'intéresse
principalement aux Angiospermes mais on retrouvera quelquefois les Gymnospermes."
•Les Spermatophytes se divisent en 2 embranchements:"
-Gymnospermes: graines exposées sur une feuille de support, il n’y a pas de
fermeture de la feuille sous la forme de carpelle. 3 sous-embranchements:
Cycadées, ginkgos et conifères "
-Anthophytes, qui regroupent:"
‣Gnétophytes: groupe intermédiaire"
‣Angiospermes: groupe le plus important au niveau végétal (450000 espèces), très
diversifié au niveau des caractéristiques végétatives, du développement végétatif
et développement floral."
!
!
!
!
1

•On observe une origine commune aux Angiospermes, qui est un groupe
monophylétique. Tout un éventail de plantes vont être retrouvées dans les
Angiospermes, mais on étudiera les classes des:"
-Eudicotylédones qui présentent les caractéristiques des plantes eudicotylédones"
-Monocotylédones ou Monocotylées, qui présentent les caractéristiques des plantes
monocotylédones."
!
!B) Germination
!!1. Rappel: cycle biologique du végétal
•La phase sporophytique est la plus importante chez les Angiospermes."
!
2

!!2. Rappel: développement embryonnaire d’un
Angiosperme Eudicotylédone
!
•Au cours du processus de la germination, la gemmule se développe et donne une tige
feuillée au-dessus des deux cotylédons. L’épicotyle sera la partie de la jeune plante en
germination située au-dessus de l'insertion des cotylédons. L’hypocotyle est la
connexion entre tissus aériens et tissus racinaires. Le radicule est un début de racines
prêtes à se développer."
!
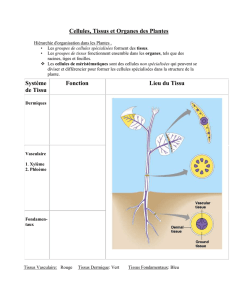
•Très peu de tissus sont formés au stade embryonnaire:"
-l’épiderme protège l'embryon de l’environnement"
-le parenchyme est un tissu de remplissage"
-Au centre, le procambium joue un rôle dans la conduction de la sève brute et de la
sève élaborée. "
•Il n’y a pas à ce stade de formation des organes qu'on retrouve chez l'organisme
adulte. Contrairement aux animaux, le développement embryonnaire permet
simplement une ébauche, l’organogénèse est post-embryonnaire et se fait grâce au
développement du méristème apical caulinaire et du méristème apical racinaire."
!
!!3. Mode de germination
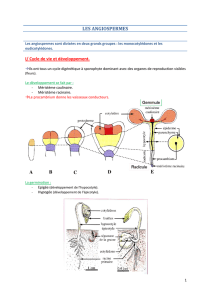
•Lorsqu’on met une graine dans un sol et que les conditions sont favorables: "
-le radicule subit un gravitropisme positif, c’est à dire une attraction vers le centre de
la Terre."
-au niveau du gemmule, la partie qui se développe subit un gravitropisme négatif et
dès qu'elle sort de la terre, la partie aérienne subit un phototropisme positif."
•Dans un premier temps, le développement se fait sous forme hétérotrophe, aux dépens
des ressources énergétiques présentes dans la graine au niveau des cotylédons ou de
l’albumen."
!
!
!
3

•On différencie 2 types de germination chez les Angiospermes:"
-germination épigée: développement de l'hypocotyle très important qui assure la
croissance et la sortie de la terre. Les cotylédons font une partie de la
photosynthèse."
-germination hypogée: c'est la partie épicotyle qui se développe, les cotylédons
restent dans le sol et n'ont aucun rôle dans la photosynthèse."
!
!C) Méristème primaire et méristème secondaire
•Une fois que la plante a développé ses différentes feuilles, elle devient autotrophe. La
plante est indépendante de la source d’énergie des réserves."
•La plante possède 2 méristèmes représentés sous forme de petites cellules
méristématiques qui ressemblent davantage à des cellules animales:"
-petites cellules"
-vacuole très limitée"
-noyau très développé"
•Ces cellules méristématiques se divisent de façon très active et se différencient
progressivement en devenant des cellules végétales fonctionnelles:"
-grandes cellules"
-vacuole très développée "
4

-noyau très réduit"
•Les méristèmes fonctionnent pendant toute la durée de vie de l'organe végétal%(autre
différence avec les cellules animales), donc jusqu'à des milliers d'années chez certains
arbres."
•Avantage du développement post-embryonnaire du monde végétal: très adaptable et
flexible par rapport aux conditions de l'environnement (sol, eau, vent...)%puisque les
plantes ont une vie fixée."
!
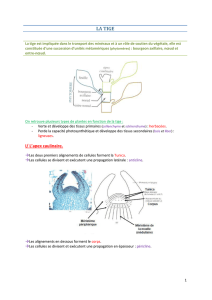
!!1. Méristèmes primaires apicaux d’élongation
!
•Le MAC et le MAR ont donc un rôle d'élongation indéfini de la plante tant qu'elle survit."
•Les méristèmes développent ici des structures primaires. Il y a a peu près le même style
de développement dans les 2 méristèmes: ce sont des structures assez désorganisées
au niveau spatial qui partent d'un point méristématique et rayonnent autour de ce point
par divisions actives de type transversal."
!
!!2. Méristèmes secondaires d’élargissement
5
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
1
/
51
100%