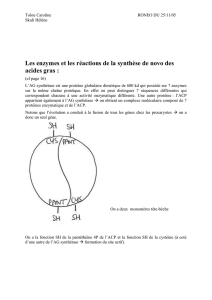
– UE :VI –

2015-2016 Métabolisme des lipides
Métabolisme des lipides
– UE :VI –
biosynthèse et catabolisme des AG
Annexe sur moodle pour les diapos
Semaine : n°7 (du 19/10/15 au
23/10/15)
Date : 20/10/2015
Heure : de 8h à 9h Professeur : Pr. Brousseau
Binôme : n°24 Correcteur : 23
Remarques du professeur (Diapos disponibles, Exercices sur le campus, Conseils, parties importantes
à retenir, etc.)
•Mettez les informations
•Sous forme
•De liste
PLAN DU COURS
PLAN DU COURS
!""
1/9

2015-2016 Métabolisme des lipides
#
–$%&
–#&% !"#$%&'
$ '()*+ +
!&
(
$ # %) *
" + ,
-%(
–$
–$'(!'.%/0 &
–# &,+ 1$2%3 4
#5$ $
–-&" $%660 7!
3&
2/9

2015-2016 Métabolisme des lipides
1$% 8 61$%#1$%
6%1$36%96$2 61$%(
$6%96%/
6 $% $% : % $% 1$%
1$%31$%(
*
: $% ; 6 %9 ' .%/0 , ; ' .%/ ; :
$%<=0;0 2;6%/
0 2 6 0 2 0 2
(
8$2 !6$2 6$%
3&(
8../0.
36%9($,>
#( 2 ? #
7%9(
$ %9 %9
@6%9>7A
!
B!131&
1 " 2! 3 4 " +5
6!"
B#+78"9
= 7 "4
4$!C& !$:& DE3
!E3 &(
3/9

2015-2016 Métabolisme des lipides
=2# " :3
F(84 !1&3!1 &(((
3 $:31
1E!GGE&
'."9
BC4%)
14(
,!79% !!1E&& 3:
;53:#53:53:<
$'4(
8
%)(
23*
! H C &( B # '
-0(
I
$!&
–2 =;
I D E!$E$7&#
J43DE
4/9

2015-2016 Métabolisme des lipides
–8=6
–I=K! /
H4-&
8 = . = 34%)
$$:(
L - ! 8 F # 4
4 ( 2 K K
$K$J%)
–8 = 9 C D K
4$ 7!
'K(
, = ;5 = 5 !5 = !
6=
2 ".7"4!&3
!DE&! 16
G6&(
216!G6&
)"%<%
15=$1 =$+
-(2151
! %)
>%)(&
5/9
 6
7
8
9
6
7
8
9
1
/
9
100%