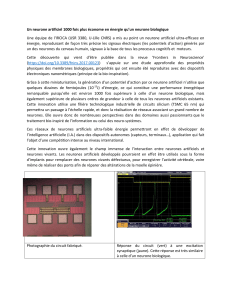
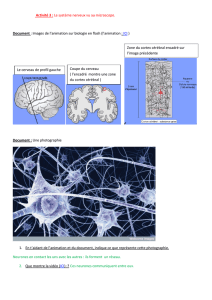
UNIVERSITÉ OUVERTE 2013-‐2014 Du neurone au cerveau: Une

!"#$%&'#()*+!$%&(%*,-./0,-.1*
Du#neurone#au#cerveau:#
Une#promenade#dans#la#complexité#
#du#système#nerveux**
23450673883*9358:*
;4<=84>=783*?4@A83*3>*
':B>AC3B*D=CEF3G3BH*
!57I38B7>J*6487B*K7L38=>*
MNEOPPQQQRCBSRT57I0E487B0L7L38=>RU8P
VM358:P*
W3450E73883RM358:XT57I0E487B0L7L38=>RU8*

D=T8B*1*
;3B*8JB34TG*L3*53T8=53BO*
D=CC35>*7FB*U4<87YT35>*L3B*E8=E87J>JB*5=TI3FF3BH**
F4*CJC=783*S3FFTF4783*
23TL7*,Z*5=I3C<83*,-./*

&JBTCJ*L3B*S=T8B*E8JSJL35>B*[.\*
• ;3*U=5S@=553C35>*LT*S38I34T*7CEF7YT3*F4*S78STF4@=5*
L]75U=8C4@=5*35>83*T5*^845L*5=C<83*L3*S3FFTF3B*
[!"#$%!"&\*
• ;]75U=8C4@=5*3B>*L]4<=8L*JF3S>87YT3H*B=TB*F4*U=8C3*
L]T53*I4^T3*L3*E=>35@3F*S78STF45>*L3ET7B*F]3G>8JC7>J*
L35L87@YT3*WTBYT]_*F4*>38C7547B=5*B:54E@YT3*
['%("!)"*+,-./)%!\H*_*F]75>J873T8*LT*53T8=53*
• ;3*E4BB4^3*L3*F]75U=8C4@=5*L]T53*S3FFTF3*_*F]4T>83*3B>*
4BBT8J*E48*F4*F7<J84@=5*L3*C=FJSTF3B*
[!"#$%($.!&0"1"#$&\H*_*>84I38B*F]3BE4S3*B:54E@YT3*
• ;4*F7<J84@=5*LT*53T8=>845BC3N3T8*[23%/4(%&"\*3B>*
LJSF35SMJ3*E48*T53*4T^C35>4@=5*LT*D4,`*L45B*F4*
>38C7547B=5*75LT7>3*E48*F]4887IJ3*LT*E=>35@3F*L]4S@=5***

&JBTCJ*L3B*S=T8B*E8JSJL35>B*[,\*
• ;3*53T8=>845BC3N3T8*F7<J8J*B3*aG3*BT8*L3B*$5/"'("#$&*
E8JB35>B*BT8*F3*53T8=53*S7<F3*
• D38>475B*8JS3E>3T8B*B=5>*L3B*S454TG*YT3*F4*F747B=5*LT*
53T8=>845BC3N3T8*=TI83*[$5/"'("#$&+6%!%($%'"&\*
• D3N3*=TI38>T83*C=L7a3*F3*E=>35@3F*L3*83E=B*LT*
53T8=53H*LJSF35SM45>*T5*E=>35@3F*L]4S@=5*[!"#$%!"+
"3/6(.("#$\*=T*I388=T7FF45>*B4*^35AB3*[!7+6!8696("#$\*
• K]4T>83B*4^7BB35>*BT8*F3*CJ>4<=F7BC3*S3FFTF4783*
[$5/"'("#$&+05(.9%($%'"&\*E48*F]75>38CJL74783*L3*
E8=>J753B*L3*B7^54F7B4@=5*[E8=>J753B*b\*
• K45B*S3*S4BH*F3B*3c3>B*B=5>*L7I38BO*C=L7aS4@=5*L3*
F]4S@I7>J*L3*E8=>J753BH*LJC4884^3*LT*E8=^84CC3*
^J5J@YT3\****

D=CC35>*F3B*53T8=53B*>84I47FF35>*35B3C<F3*
• D=CC35>*T53*d*CJC=783*e*E3T>*4EE484f>83*
SM3g*T5*75I38>J<8J*
• %>*SM3g*5=TBh*
• ;3*S38I34T*IT*S=CC3*T5*8JB34T*L3*53T8=53B*
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
1
/
70
100%