Open access

! ! !!
! !
Université de Caen Basse-Normandie
Ecole doctorale Normande de Biologie Intégrative, Santé et Environnement
Thèse de doctorat
présentée et soutenue le : 23/10/2013
par
Ingrid WALDSCHMIDT
pour obtenir le
Doctorat de l’Université de Caen Basse-Normandie
Spécialité : Physiologie, Biologie des organismes
préparée dans le cadre d’une cotutelle internationale de thèse
entre l’Université de Caen Basse-Normandie et l’Université de Liège
Effet de l’effort, l’entrainement et l’inflammation sur l’immunité innée des
voies respiratoires profondes chez le Trotteur Français
Directeurs de thèse : Audigié Fabrice (Université de Caen)
et Art Tatiana (Université de Liège)
Jury
Cadoré Jean-Luc, Professeur des Universités, Ecole Nationale Vétérinaire de Lyon (rapporteur)
Kirschvink Nathalie, Professeur des Universités, Université de Namur (rapporteur)
Vabret Astrid, Professeur des Universités, Université de Caen
Pirottin Dimitri, chercheur, Université de Liège
Dupuis-Tricaud Marie-Capucine, chercheur, Ecole Nationale Vétérinaire d’Alfort
Audigié Fabrice, Professeur des Universités, Ecole Nationale Vétérinaire d’Alfort (directeur de
thèse)
Art Tatiana, Professeur des Universités, Université de Liège (directeur de thèse)


REMERCIEMENTS
Je souhaiterais remercier mes encadrants de thèse, Fabrice Audigié, pour avoir accepté
d’encadrer administrativement cette thèse, et plus particulièrement Tania Art pour son
accueil, son encadrement scientifique, ses conseils pertinents et sa bonne humeur
permanente ! Merci à tous les membres du jury, pour avoir accepté mon invitation.
A Marie-Capucine Dupuis-Tricaud, pour son accueil chaleureux à mon arrivée en Normandie,
son aide précieuse et sa disponibilité tout au long du projet de recherche ainsi que pour tous
les agréables moments partagés au CIRALE ou sur la plage de Cabourg !
Aux membres du CIRALE : au Pr Jean-Marie Denoix, pour m’avoir accueilli, à l’équipe de
Médecine Sportive : Julien (pour tes fameuses « julienades » et l’entrainement non sans
difficulté des chevaux expé), puis Elodie (à nos critiques sans fin du climat normand !),
Thibault, Dagmar, à l’équipe de pathologie locomotrice : Sandrine, Virginie, Lélia (vous vous
souviendrez longtemps des péripéties de nos chevaux !) et toute l’équipe technique : Aurélie,
Marie-Mélisande, Florence, Marie, Manu, Alain, Quentin.
A l’équipe du CEMESPO : Linda, Irène, Sophie pour nos moments de galères expérimentales
partagés, bonne chance pour la suite, ça arrive plus vite qu’on ne le pense ! A l’équipe
technique : Raja, Ilham, pour votre efficacité et votre bonne humeur, Jean-Clément, pour les
grignotages autour de la brouette à granulés, aux anciennes du CEMESPO : Eve, Audrey,
Clémence et Céline pour nos balades équestres dans les bois du Sart ! Et enfin au Pr Pierre
Lekeux qui dirige toute cette joyeuse troupe. C’était toujours un plaisir de venir travailler à
Liège.
A l’équipe d’immunologie du GIGA et plus particulièrement Dimitri, pour son encadrement
technique et sa disponibilité, Fabrice et Christophe pour leurs conseils avisés, ainsi que tous
les autres membres, pour les sympathiques moments partagés à Durbuy.
A l’équipe du laboratoire Frank Duncombe : Guillaume, Pierre-Hugues, Stéphane, Eric,
Albertine, Loïc, Sophie, Christine, Erika, etc, pour votre accueil au sein de votre service et
nos réflexions scientifiques entre la Normandie et Liège.

A l’équipe de l’ANSES pour leur accueil toujours chaleureux.
Aux membres de la fondation HIPPOLIA, pour leur investissement dans la recherche équine.
Aux vétérinaires et propriétaires qui ont participé à la partie sur les chevaux de terrain.
Aux co-financeurs qui ont permis l’aboutissement de ce projet : l’Institut Français du Cheval
et de l’Equitation et la Région Basse-Normandie.
Aux chevaux qui ont eu la patience de subir les prélèvements tout au long du protocole, à
leurs péripéties quasi hebdomadaires. A Rita, Rosie, Une, Ulga, Véra, Voyou, Vaillant,
Vigilance, Udar et plus particulièrement Unika !
Enfin, le principal, merci à ma famille et amis, Vincent, pour m’avoir supportée durant ces
rudes années normandes, mon tonton, mon grand-père et surtout ma maman pour m’avoir
toujours poussée plus haut.

TABLE DES MATIERES
.......................................................................................................................... 1!LISTE DES FIGURES
...................................................................................................................... 4!LISTE DES TABLEAUX
............................................................................................................. 5!LISTE DES ABREVIATIONS
................................................................................................................................................ 8!RESUME
.......................................................................................................................................... 10!SUMMARY
........................................................................................................ 12!INTRODUCTION GENERALE
............................................................................................................. 14!REVUE DE LITTERATURE
1!AFFECTIONS RESPIRATOIRES PROFONDES DU JEUNE CHEVAL ATHLETE ET IMMUNITE
INNEE : ................................................................................................................................. 15!
1.1!Infections virales : une réponse immunitaire insuffisante ? .......................... 15!
1.2!Maladie inflammatoire des voies respiratoires : une réponse immunitaire
excessive ? ....................................................................................................................... 17!
1.3!Mécanismes de défense pulmonaire, focus sur l’immunité innée : ............... 20!
1.3.1!Mécanismes passifs : ...................................................................................... 20!
1.3.2!Acteurs cellulaires : ........................................................................................ 21!
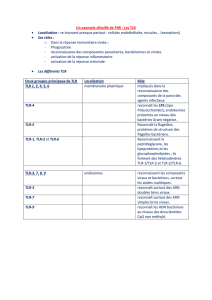
1.3.3!Reconnaissance des pathogènes et réponse immunitaire innée : ............... 24!
a)!Structure des TLR : .......................................................................................... 25!
b)!TLR et ligands : ................................................................................................ 26!
c)!Activation des TLR et transduction du signal pathogène : .............................. 28!
d)!Interface avec l’immunité acquise : ................................................................. 35!
2!IMMUNITE INNEE, EXERCICE ET ENTRAINEMENT : .................................................... 38!
2.1!Etudes sur l’Homme : ........................................................................................ 38!
2.1.1!Données épidémiologiques : .......................................................................... 38!
2.1.2!Données expérimentales : .............................................................................. 39!
a)!Exercice ponctuel : .......................................................................................... 39!
b)!Entrainement : ................................................................................................. 42!
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
147
148
149
150
151
152
153
154
155
156
157
158
159
160
161
162
163
164
165
166
167
168
169
170
171
172
173
174
175
176
177
178
179
180
181
182
183
184
185
186
187
188
189
190
191
192
193
194
195
196
197
198
199
200
201
202
203
204
205
206
207
1
/
207
100%