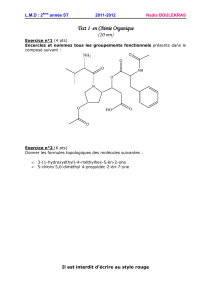
Corrigé ET1_2010

BIO241 Examen terminal 1ère session (25 mai 2010)
Documents et calculatrices non autorisés
ATTENTION : Les parties A, B et C doivent être traitées sur des copies
séparées
Notation sur 100 points
PARTIE A
Cours de Mme C. Breton (44 points)
Question 1 : les MPT 12 pts
Les mécanismes de régulation de la fonction des protéines impliquent souvent
des modifications post-traductionnelles (MPTs).
1/ Quelles sont les 2 grandes catégories de MPTs ? 2 pts
- addition covalente de groupements plus ou moins gros
- clivage de la chaîne polypeptidique
2/ Dans le cas d’une modification par phosphorylation : 8,5 pts
a. Quels sont les résidus d’acides aminés concernés par cette modification
Ser, Thr, Tyr
b. Donner la formule semi-développée d’un résidu d’acide aminé (un exemple
au choix) avant et après modification
1 pt pour la sérine non modifiée
2 pts pour la serine modifiée (avec bonne indication de la charge)
c. S’agissant d’une modification réversible, donner le nom des enzymes
impliquées dans les réactions de phosphorylation et dé-phosphorylation
Protéines kinases et phosphatases
d. Quelle est l’incidence de cette modification sur la structure de la protéine
Introduction d’une charge négative entrainant une modification de
l’environnement protéique. Au niveau des enzymes, cela se traduit le plus
souvent par un effet d’activation/inactivation de la protéine (On /Off).

3/ Donner trois autres exemples de MPTs 1,5 pts
Réactions d’acylation : ex N-acétylation, N-myristoylation, S-
palmitoylation, ubiquitination,
Réactions d’alkylation : ex méthylation
Autres exemples : glycosylation, maturation protéolytique, hydroxylation,
biotinylation
Question 2 : Hémoglobine 6 pts
Les mobilités électrophorétiques à pH 8,6 d’une série de molécules
d’hémoglobine normale et mutantes sont données ci-dessous :
électrode - l l l l l électrode +
a b Hb c d
normale
i k h m
Faire correspondre les positions a, b, c et d avec les molécules Hb mutantes
indiquées ci-dessous :
Hbh : glycine remplacée par aspartate en position β69
Hbi : glutamate remplacé par lysine en position β6
Hbk : asparagine remplacée par lysine en position α68
Hbm : lysine remplacée par glutamate en position β95
Justifier votre réponse
Réponse : a=Hbi b=Hbk c=Hbh d=Hbm
Raisonnement : il fallait évaluer quelle était l’influence de la mutation sur la
charge nette de la protéine par rapport à l’Hb normale
Hbh : -1
Hbi : +2
Hbk : +1
Hbm : -2
Question 3 : Glycolyse 20 pts
1. Certaines réactions de la glycolyse se produisent dans des conditions
d’irréversibilité thermodynamique. Indiquer lesquelles.
Ecrire en détail les réactions (noms et formules semi-développées des réactants
et noms des enzymes) (6 pts)
1. Glucose + ATP Glucose-6-P + ADP hexokinase (HK)
3. Fructose-6-P + ATP Fructose-1,6-bis-P + ADP Phosphofructokinase (PFK)
10. Phosphoenolpyruvate + ADP Pyruvate (Pyr) + ATP Pyruvate kinase (PK)
2. Chez l’homme, en condition anaérobie, le pyruvate subit une transformation.
(7 pts)

a. Donner le nom de cette transformation. Ecrire en détail la ou les
réaction(s) (noms et formules semi-développées des réactants et noms des
enzymes)
Fermentation lactique
Pyruvate + NADH,H+ lactate + NAD+ Lactate deshydrogenase (LDH)
b. Dans quel(s) type(s) cellulaire(s) ou tissu(s) cette réaction a-t-elle lieu ?
muscle squelettique (en activité) et GR
c. Quel est l’intérêt principal de cette réaction ?
régénérer le NAD+
3. Dans le contexte de la glycolyse, quel est le point commun entre ATP,
Fru2,6-bisP et citrate ? (3 pts)
Ce sont des molécules qui vont moduler la vitesse de glycolyse. Ce sont plus
précisément des inhibiteurs (ATP et citrate) ou des activateurs (F2,6-bisP)
allostériques de la PFK
4. Lors d’un petit déjeuner, il est ingéré 17g de saccharose. Calculer le
nombre de moles ATP formées (en conditions aérobies) en supposant que
les monosaccharides libérés à partir du saccharose (glucose et fructose)
empruntent immédiatement la voie de la glycolyse. Vous utiliserez les
valeurs hautes de P/O.
MM saccharose : 340 g.mol-1 (4 pts)
Une mole de saccharose libère une mole de glucose et une mole de
fructose. Chacun de ces sucres permet de former 38 ATP, soit 76 moles
ATP au total/ mole de saccharose.
17/342= 0,05 moles de saccharose
0.05x76 = 3,8 moles ATP
Question 4 : Glycogène 3 pts
1/ Donner la structure du glycogène et son rôle dans l’organisme.
Polymère de résidus glucose liés en 1,4. Branchement en 1,6.
Forme stockage du glucose rapidement mobilisable
2/ Indiquer les lieux de stockage du glycogène dans l’organisme.
Foie et muscles
3/ Quelles sont les deux enzymes clé du métabolisme du glycogène (synthèse et
dégradation) ?
Glycogène synthase et Glycogène phosphorylase

Question 5 : ATP synthase 3 pts
L’ATP synthase est un complexe protéique enzymatique qui permet de produire
de l’ATP à partir d’ADP et de Pi. Indiquer sa localisation cellulaire et quelle est la
source directe d’énergie utilisée pour cette synthèse ?
Localisation : membrane interne mitochondriale
L’énergie provient d’un gradient de protons qui s’établit au niveau de la
membrane interne de la mitochondrie. Il s’agit d’un couplage entre un
transport spontané de protons (entre espace intermembranaire et la
matrice) et une réaction chimique non spontanée.
************************
PARTIE B
Bioénergétique et Enzymologie : cours de Mme F. Cornillon
Les hydrogénases sont des enzymes qui catalysent la réaction réversible
d'oxydation de l'hydrogène moléculaire selon la réaction suivante :
E°'= -0,42V
Une hydrogénase a été purifiée à partir d'une bactérie sulfato-réductrice. Une
analyse par ultracentrifugation analytique indique que la protéine native a un
poids moléculaire de 50 kDa.
L'analyse par gel d'électrophorèse en présence de SDS donne le résultat suivant
présenté sur la figure 1 :
Figure 1 :
électrophorèse sur gel de
polyacrylamide en présence de SDS
(dodécyl sulfate de sodium)
A : Marqueurs de poids moléculaires
suivants : 10 KDa, 20 KDa, 30 KDa,
50 KDa et 60 Kda.
B : Echantillon d'hydrogénase
purifiée

Question 1 : 6 pts
1/ Rappeler le principe de l'électrophorèse sur gel de polyacrylamide en présence
de SDS.
L'application d'un champs électrique permet de faire migrer les molécules
chargées. Le gel se comporte comme un tamis moléculaire : il retarde les
grosses molécules et laisse passer entre ses mailles les plus petites.
Le SDS, sodium dodecyl sulfate, est un détergent anionique : il dénature les
protéines (rupture des liaisons faibles uniquement) en les tapissant de charges
négatives.
Ainsi, en présence de SDS, l'électrophorèse permet de séparer les protéines
uniquement selon leur masse molaire. En effet, les protéines sont dénaturées et
chargées uniformément négativement par le SDS : elles migrent toutes vers
l'anode.
2/ Que pouvez-vous conclure de l'expérience précédente ?
L'hydrogénase est formée de deux sous unités de 40 et 10kDa respectivement,
reliées par des liaisons faibles.
Question 2 : 9 pts
Les électrons produits lors de la réaction d'oxydation de l'hydrogène moléculaire
sont transférés par l'hydrogénase à un accepteur d'électrons.
Afin de pouvoir suivre la réaction de consommation de l'hydrogène par
l'hydrogénase, nous utilisons un composé coloré : le méthyl viologène (MV2+). La
réaction de réduction du méthyl viologène est la suivante :
E°'=-0,45V
Le méthyl viologène réduit (MV+) absorbe la lumière à 600 nm, ce qui lui confère
sa couleur violette, alors que la forme oxydée (MV2+) est incolore.
1/ Ecrire l'équation bilan de réduction du méthyl viologène par l'hydrogène
moléculaire catalysée par l'hydrogénase.
H2 + 2 MV2+ 2H+ + 2MV+
2/ Calculer le potentiel d'oxydoréduction dans les conditions standard apparentes
de la réaction bilan de réduction du méthyl viologène par l'hydrogène
moléculaire.
ΔE°' = E°'MV2+/MV+ - E°'H+/H2 = -0,45 -(-0,42) = -0,03V
3/ Cette réaction de réduction du méthyl viologène par l'hydrogène moléculaire
est-elle spontanée dans les conditions standard apparentes ? Justifier.
ΔG°' = - n F ΔE°' = -2x96500x(-0,03) >0 donc réaction non spontanée dans les
conditions standard apparentes
4/ Pourrez-vous faire l'étude cinétique de cette réaction, pour mesurer l'activité
de l'hydrogénase, indifféremment dans le sens 1 ou 2 ? Justifier.
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%