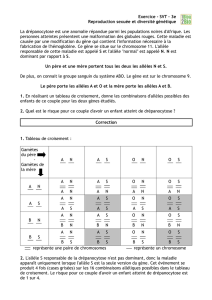
IV. Les autres rapports alléliques

GÉNÉTIQUE
Chapitre 2 :
Transmission des caractères

SOMMAIRE
I. Définitions
1. Gène-Locus-Allèle
a. Gène
b. Locus
c. Allèle
2. Génotype-Phénotype-
Caractère
a. Génotype
b. Phénotype
c. Caractère
3. Homozygote-Hétérozygote-
Hybride
a. Homozygote-
Hétérozygote
b. Hybride
4. Gènes liés-Dominance-
Croisement
a. Gènes liés
b. Dominance
c. Croisement
5. Polymérie-Pléiotropie
6. Symboles
2. Dominance incomplète
a. Rappel
b. Exemple
3. Gène létal
a. Définition
b. Exemple
4. Pénétrance
a. Définition
b. Exemple
5. Allèles multiples
a. Définition
b. Exemple
V. Dihybridisme
1. Définition
a. Définition
b. Exemple
2. Test-cross
a. Objectif
b. Exemple
3. Rapport dihybrides modifiés
a. Définition
b. Exemple
VI. Polyhybridisme
II. Les lois de Mendel
1. Gregor Mendel
2. Matériel et méthode utilisés par
Mendel
3. Les lois de Mendel
a. La première loi
b. La deuxième loi
c. La troisième loi
III. Monohybridisme
1. Définition
a. Définition
b. Exemple
2. Le test-cross
a. Définition
b. Exemple
3. Le back-cross
a. Définition
b. Exemple
IV. Les autres rapports alléliques
1. Allèles codominants
a. Rappel
b. Exemple

Le terme de gène est tellement large qu’il
est difficile d’en donner une définition
Un gène peut néanmoins se définir comme
un enchaînement de bases azotées, une
séquence d’ADN, destiné à être transcrit en
ARN
Il commence généralement par le
promoteur dont le rôle est de permettre
l’initiation et la régulation de la transcription,
et se termine par une séquence de
terminaison
La molécule d’ARN transcrite peut être soit
traduite en une chaîne polypeptidique, soit
directement fonctionnelle
1. Gène-Locus-Allèle a. Gène
3
I. DÉFINITIONS

1. Gène-Locus-Allèle a. Gène
Le gène correspond à une unité de
fonction: c’est la plus petite portion de
matériel génétique nécessaire et
suffisante à la réalisation d’une fonction
donnée
C’est une unité d’hérédité contrôlant
un caractère particulier
4
I. DÉFINITIONS

Le locus est l’emplacement précis d’un
gène sur un chromosome
La localisation biochimique d’un gène est
fondée sur un modèle standard de bandes
claires et sombres obtenues après
application d’une technique de coloration
Le gène est d’abord localisé par le
numéro du chromosome pour les
chromosomes non sexuels (1 à 22)et par
une lettre (X ou Y) pour les chromosomes
sexuels
Une lettre suit la désignation du
chromosome, p (désignant le petit bras du
chromosome) ou q (le grand bras du
chromosome)
1. Gène-Locus-Allèle b. Locus
5
I. DÉFINITIONS
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
1
/
63
100%