Organisation du vivant Végétal Développement et plans d`organisation

Organisation du vivant Végétal
Développement et plans d'organisation
I] Introduction aux Angiospermes
Partagent un ancêtre avec les gymnospermes
(groupe monophylétique).
Chez les angiospermes, on retrouve des formes arborescentes (palmier
par exemple). Les angiospermes sont divisés en deux grand groupes :
les monocotylédones et les eudicotylédones.
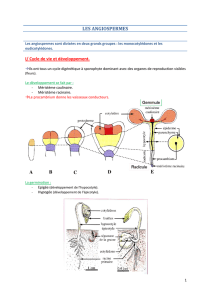
1) Cycle de vie et développement
Les angiospermes ont un cycle digénétique à sporophyte
dominant, avec des organes de reproduction visibles (fleurs).
Après fécondation, le développement
embryonnaire se fait par deux pôles
méristématiques : méristème caulinaire, et
méristème racinaire. L'embryon est une
ébauche de la plante adulte, mais seulement
au niveau des tissus : aucun organe n'est
identifiable. Le procambrium donnera les
vaisseaux conducteurs.
La germination peut être soit épigée (développement de l'hypocotyle,
donc sortie de la « graine » du sol), ou hypogée (développement de
l'épicotyle, donc la graine reste dans le sol).
La croissance de la plante est dirigée en partie par les forces de
gravité : c'est le gravitropisme. On le dit positif au niveau des racines
(cherche la gravité), et négatif au niveau du méristème caulinaire
(contre la gravité).
La croissance est également régie par le phototropisme (dépendant
des photons) : positif au niveau du méristème caulinaire (cherche la
lumière) et négatif au niveau des racines (fuit la lumière).
Au début, la graine n'est pas photosynthétique. Elle utilise ses
ressources accumulées pour débuter sa croissance, puis dès qu'elle a
des feuille, passe à la photosynthèse.

2) Les méristèmes
Les cellules méristématiques sont totipotentes, et se divise souvent par mitose. Elles
vont former les organes : c'est l’ontogenèse, et des cellules histologiques
spécifiques : c'est l'histogenèse. Ces deux mécanisme permette une certaine
adaptabilité de la plante, qui pourra différencier ses tissus et organes en fonction de
l'environnement. De plus, les méristèmes vont être responsables de l'élongation de
la plante : tout au long de la vie de la plante, les méristèmes apicaux (caulinaires et
racinaires) vont s'éloigner.
Les méristèmes primaires sont des petits centres de développement peu organisés
principaux. Les méristèmes secondaires apparaissent plus tard dans le
développement du végétal (méristèmes néoformés) et ont un rôle dans
l’épaississement du végétal. Les méristèmes secondaires apparaissent que dans
certains types de plantes (plutôt les plantes pérennes), et seulement chez les
eudicotylédones. Les méristèmes secondaires sont, eux, plus organisés.
Le but des méristème est de générer un grand nombre de cellules par division méiotique (mérèse), qui vont ensuite grandir
(auxèse) et se différencier. Pendant l'auxèse, la paroi est flexible et peut croitre, puis se rigidifie quand la taille finale est
atteinte. Cette rigidité permet de résister à la pression de turgescence, qui permet notamment le maintient de la plante.
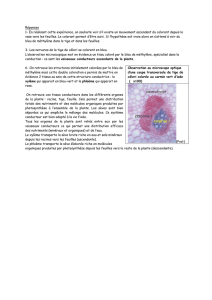
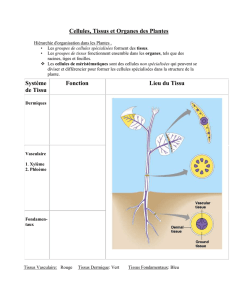
3) Différenciation cellulaire et histogenèse des tissus primaires
La différenciation peut mener à la mort de la
cellule, qui sera ensuite utilisé pour faire les
vaisseaux conducteurs. Les méristèmes
génèrent des cellules indifférenciées
organisées en fils cellulaires (procambrium).
Dans le cas du xylème, les cellules en fils
sont mise en contact les unes aux autres par
perforation de la paroi. La paroi secondaire
s'épaissit et se perfore par activité
enzymatique pour former les ponctuations
(échanges d'eau et de minéraux), le tonoplaste
(membrane vacuolaire) et le noyaux
dégénèrent ensuite.
Issu également du tissu du
procambrium. Les paroi cellulosique
reste intactes, mais les
plasmodesmes sont agrandis, le
noyau et la vacuole dégénèrent. Ces
cellules énucléées sont associées à
des cellules compagnes qui se
trouvent à proximité, et qui
fournissent aux cellules du phloème
les nutriments essentiels. Si une
cellule du phloème est touchée, une
cellule compagne peut se diviser
pour reformer la cellule manquante.

Cependant, la différenciation cellulaire est réversible. Toutes les cellules
végétales sont capables de se dédifférencier pour revenir dans un état
méristématiques. A partir de ce méristème, elles vont régénérer la structure
manquante. Cependant, ces capacités ne sont pas anarchiques, mais sous
contrôle de phyto-hormones, ainsi qu'un contrôle sur la position des
cellules. De plus, les processus de différenciation sont également soumis à
un contrôle important.
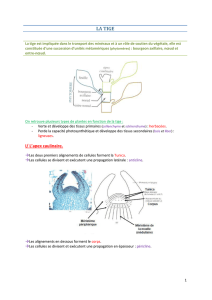
Il y a deux grand types de tissus :
•Tissus de protection qui dérivent du
protoderme
•Tissus conducteurs qui dérivent du
procambrium
Au niveau des feuilles, une couche de cuticule se
dépose sur l'épiderme (tissu de protection) pour
empêcher les pertes d'eau, laissant au stomate
l'entière régulation hydrique.
Les tissus vasculaires sont issus de deux pôles du
procambrium, l'un criblé (génère le phloème),
l'autre vasculaire qui génèrera le xylème.
Le tissus de « remplissage » (entre épiderme et
tissus vasculaires) s'appelle le parenchyme et a un
rôle dans les échanges liquides et gazeux avec les
méats.
Les collenchymes et sclérenchymes ont un rôle
dans la rigidité du végétal. Les cellules sont toutes
jointives (sans méats), avec épaississement assez
fort de la paroi (et irrégulier pour combler les
méats). Le soutien n'est pas totalement rigide, mais
permet le maintient du végétal. Pour le
schlérenchyme, les cellules sont rondes et sous
forme fibreuse, et on une paroi très épaissie (paroi
secondaire), et des dépôts de subérine et lignine
dans ces parois renforcent le soutient (résistance à
la compression).
On peut aussi trouver des tissus
sécréteurs (divers substances
attractives ou répulsives par
exemple), des canaux sécréteurs de
résines (lutte contre les nuisibles).
Les tissus racinaires sont également
un peu différent, spécialisés dans les
flux d'eau et de nutriments vers les
pôles vasculaires (endoderme et
assise génératrice de cellule
méristématiques capable de générer
méristème, et méristèmes
secondaires)
Les méristèmes primaires qui assurent l'élongation de la
plante se divisent transversalement.
Les méristèmes secondaires assurent une croissance en
épaisseurs, et génèrent de typent de tissus différents :
•Assise libéro-ligneuse (ou cambium) : Composé
de bois et liber qui vont composer le xylème. C'est
un tissu de conduction
•Assise subéro-phellodermique (phellogène) :
Composé de suber et phelloderme qui vont
composer le phloème. C'est un tissu de revêtement.

Lors de la croissance de la plante, les tissus primaires issus du méristème primaire vont
rapidement régresser (une ou deux années) au profit de tissus secondaires.
4) La racine
Lieu de synthèse de certains composés (nicotine par exemple, phyto-hormones, acide abscissique...), elles sont orientés par le
gravitropisme et fixent la plante dans le substrat.
a) Construction de l'appareil racinaire
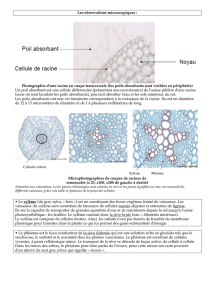
La coiffe protège l'extrémité de la racine, la zone lisse n'ayant pas de poil d'absorption, la
zone pilifère qui va assurer les transferts hydriques grâce à des poils absorbant (durée de
vie courte) et la zone subéreuses qui empêche les échanges d'eau.
La coiffe protège l'extrémité de la racine. Les cellules de la coiffe sécrètent un
mucilage qui permet d'améliorer la pénétration de la racine dans le sol. En 1, on
retrouve des organes sensoriels de la gravité, présentant des grains d'amidons,
permettant de générer un gravitropisme positif des racines en cours de
développement. En 3, sous la coiffe, on retrouve la pointe du méristème
racinaire, avec en son centre une zone quiescente. Depuis la zone
méristématique, on peut différencier trois types de tissus : le procambrium qui
va jouer un rôle dans la mise en place des cellules de la sève, le méristème
fondamental qui va générer des tissus sous forme de fils qui entourent la zone
vasculaires, et le protoderme jouant un rôle de protection.
Une fois que les cellules se sont multipliées, il y a différenciation et élongations
de ces dernières dans la zone lisse. Le premier tissu se développant est le
phloème (pour irriguer la racine en sucres), le suivant sera la xylème pour
pouvoir approvisionner le reste de la plante en sels et eau.
La plus grande élongation se fait juste au dessus de l'apex, puis les cellules
cessent de se diviser pour s'allonger et se différencier seulement.
b) Structure primaire de la racine
La disproportion entre la stèle et l'écorce est caractéristique des racines de
d'eudicotylédone. Ici on observe donc un eudicotylédone.
Le rhizoderme (ou assise pilifère) est subérifié et commence à disparaître. Il
est composé d'une unique assise cellulaire.
L'exoderme a une assise composée de 3 cellules cellulosiques. Ce tissu sera
rapidement subérifié pour empêcher les flux d'eau.
Le parenchyme cortical est un un tissu de remplissage chaotique, avec des
méats pour le transports de gaz, et peut stocker des nutriments.
L'endoderme sera spécifique en fonction de la nature de la plante. Ici on est
chez un dicotylédone. Il délimite intérieurement la partie corticale, est
imprégné de ligno-subérine (empêche les transports d'eau) et forme des
cadre de Caspary.
L'assise du péricycle est
monocouche.
Les amas de cellules
cellulosiques sous le péricycle
forment le phloème
Xylème et métaxylème sont
subérifiés (donc rose).
Protoxylème
Métaxylème

Chez les monocotylédones, la stèle est beaucoup plus grosse. Les types cellulaires sont
globalement les même, cette fois, le rhizoderme n'a que deux strates.
L'endoderme est épaissi et formé en
« U » (ou fer à cheval). Il est de plus
très hydrophobe (ligno-subérine), et
possède donc des cellules dites « de
passage » qui sont perméable à l'eau.
Dans ce système, il n'y a plus que trois
alternance xylème/phloème, mais de
très nombreuses alternance (permis
par la taille de la stèle).
On trouve également des parenchymes
médullaire central, et un parenchyme
interfascicullaire.
COMPARATIF
c) Épaississement secondaire
Au niveau de la stèle, il y a division des cellules libéro-
ligneuse (en forme d'arc) qui vont séparer xylème et
phloème. C'est la mise en place du cambium, qui va
pousser le xylème libéro-lignifié vers l'intérieur
(centripète), et le phloème subéro-phellodermé vers
l'extérieur (centrifuge).
L'épiderme est remplacé par du suber (externe) pour
former ce que l'on appelle communément le liège ainsi
que le phelloderme (interne). Ces tissus sont mis en
place par le phellogène, lui même issu du péricycle.
La plante s'élargit et contient plus que des tissus
secondaires (reliquats de tissus primaires).
Élimination de l'épiderme
et du cylindre, y compris
l'endoderme
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%