Biologie Générale Fascicule I Fichier

BIOLOGIE GENERALE
Bachelier en Sciences Biologiques
Bachelier en Sciences Chimiques
Première année
Fascicule I
Eléments de Biochimie
DAVID C. GILLAN
Année académique 2016 - 2017

2
Table des matières
1.#Rappels#de#chimie#.......................................................................................................................#4!
1.1.#Le#tableau#de#Mendeleïev#..............................................................................................................#4!
1.2.#Quantité#de#matière#.........................................................................................................................#5!
1.3.#Isotopes#................................................................................................................................................#7!
1.4.#Les#niveaux#énergétiques#..............................................................................................................#7!
1.5.#Les#orbitales#atomiques#.................................................................................................................#8!
1.6.#Formation#de#molécules#et#de#composés#ioniques.#.............................................................#10!
1.7.#La#théorie#des#orbitales#moléculaires#(MO#theory)#............................................................#11!
1.8.#La#théorie#de#l’hybridation#des#orbitales#atomiques#.........................................................#16!
1.9.#Forme#moléculaire#et#fonction#biologique#.............................................................................#19!
1.10.#Electronégativité#(
χ
)#...................................................................................................................#19!
1.11.#Les#liaisons#covalentes#...............................................................................................................#20!
1.12.#Les#liaisons#ioniques#...................................................................................................................#20!
1.13.#Les#liaisons#chimiques#faibles#..................................................................................................#21!
1.14.#Les#réactions#chimiques#.............................................................................................................#22!
2.#La#molécule#d'eau#....................................................................................................................#22!
2.1.#Structure,#liaisons#hydrogène#et#chaleur#spécifique#..........................................................#22!
2.2.#Dissociation#des#molécules#d’eau#et#pH#..................................................................................#24!
2.3.#Importance#du#pH#en#biologie#:#..................................................................................................#25!
3.#Le#carbone#et#les#hydrocarbures#........................................................................................#27!
3.1.#Exemples#de#molécules#comportant#1#atome#de#C.#.............................................................#27!
3.2.#Exemples#de#molécules#comportant#2#atome#de#C.#.............................................................#27!
3.3.#Les#hydrocarbures#..........................................................................................................................#28!
3.4.#Les#isomères#.....................................................................................................................................#31!
4.#Les#groupements#fonctionnels#............................................................................................#35!
4.1.#Le#groupement#hydroxyle#............................................................................................................#35!
4.2.#Le#groupement#carbonyle#............................................................................................................#36!
4.3.#Le#groupement#carboxyle#............................................................................................................#36!
4.4.#Le#groupement#amine#...................................................................................................................#37!
4.5.#Le#groupement#thiol#ou#sulfhydryle#.........................................................................................#38!
4.6.#Le#groupement#phosphate#...........................................................................................................#38!
5.#Les#macromolécules#organiques#........................................................................................#39!
5.1.#Les#glucides#monosaccharides#...................................................................................................#40!
5.2.#Les#dérivés#des#glucides#monosaccharides#............................................................................#55!
"#$#%#!&'!()*+,'!-./0%$1"2#!##################################################################################################################################!""!
"#$#$#!&3!45)*+,3678'!-./0%9:1"2#!##################################################################################################################!"/!
"#$#9#!&'!:;3*<=>5;α;?;45)*+,3678'!-@5*:3*!+)!α;:A@2#!#####################################################################!"/!
"#$#B#!&C3*7D'!:;3*<=>5;β;?;6)E367F)'!-β;:AG2#!###################################################################################!"H!
"#$#"#!&',!3*7D',!)E+87F)',#!###############################################################################################################################!"I!
"#$#/#!&',!3*7D',!,7357F)',#!##################################################################################################################################!/J!
5.3.#Les#diholosides#et#triholosides#...................................................................................................#61!
5.4.#Les#polysaccharides#.......................................................................................................................#66!
"#B#%#!A6>5+,'!##########################################################################################################################################################!//!
"#B#B#!&'!45>*+4K8'!#################################################################################################################################################!/I!
"#B#"#!&C78)578'!#########################################################################################################################################################!/I!
"#B#/#!&3!*'55)5+,'!###################################################################################################################################################!/I!
"#B#L#!&',!45>*+,3678+45>*38',!-@A@,2!+)!6)*+M+5>,3**N3E7D',!###################################################!H%!
5.5.#Les#lipides#..........................................................................................................................................#74!

3
"#"#%#!&',!57M7D',!D'!,=+*O34'#!###########################################################################################################################!HB!
"#"#$#!&',!57M7D',!D'!,=E)*=)E'!6'6PE3837E',#!##########################################################################################!HI!
5.6.#Les#dérivés#des#lipides#..................................................................................................................#91!
"#/#%#!&',!D<E7Q<,!D',!3*7D',!4E3,!####################################################################################################################!L%!
"#/#$#!&',!D<E7Q<,!D',!,=<E+RD',#!#######################################################################################################################!LB!
5.7.#Les#terpènes#......................................................................................................................................#97!
"#H#%#!&',!6+8+='EMK8',#!####################################################################################################################################!LH!
"#H#$#!&',!,',F)7='EMK8',#!###################################################################################################################################!LI!
"#H#9#!&',!D7='EMK8',#!############################################################################################################################################!LL!
"#H#B#!&',!=E7='EMK8',#!########################################################################################################################################!%J%!
"#H#"#!&',!=<=E3='EMK8',#!###################################################################################################################################!%J$!
5.8.#Les#protéines#ou#polypeptides.#...............................................................................................#103!
"#I#%#!&',!$J!3678+3*7D',#!###############################################################################################################################!%J9!
"#I#$#!&',!3678+3*7D',!'=!5C3576'8=3=7+8!N)6378'#!###############################################################################!%%%!
"#I#9#!&',!3678+3*7D',!E3E',#!#########################################################################################################################!%%%!
"#I#B#!&',!3678+3*7D',!6+D7(7<,!3)!,'78!D'!53!ME+=<78'#!#####################################################################!%%$!
"#I#"#!&',!3678+3*7D',!8+8!3,,+*7<,!S!D',!ME+=<78',#!#########################################################################!%%$!
"#I#/#!TE78*7M35',!ME+ME7<=<,!MN>,7F)',!D',!3678+3*7D',#!################################################################!%%$!
"#I#H#!&3!5737,+8!M'M=7D7F)'#!############################################################################################################################!%%B!
"#I#I#!&',!M+5>M'M=7D',!'=!5')E!,<F)'8U34'#!#############################################################################################!%%"!
"#I#L#!&3!,=E)*=)E'!=E7D76'8,7+88'55'!D',!ME+=<78',!############################################################################!%$J!
"#I#%J#!.53,,7(7*3=7+8!D',!ME+=<78',#!############################################################################################################!%$/!
5.9.#Les#acides#nucléiques#.................................................................................................................#130!
"#L#%#!&',!8)*5<+=7D',!########################################################################################################################################!%9J!
"#L#$#!&',!8)*5<+,7D',#!#######################################################################################################################################!%9B!
"#L#9#!&3!,=E)*=)E'!ME7637E'!D',!3*7D',!8)*5<7F)',#!#############################################################################!%9I!
"#L#B#!&3!,=E)*=)E'!,'*+8D37E'!D',!3*7D',!8)*5<7F)',#!#########################################################################!%BJ!
6.#Références#..........................................................................#VEE')E!W!&'!,748'=!8X',=!M3,!D<(787#!

4
1.#Rappels#de#chimie#
1.1.#Le#tableau#de#Mendeleïev#
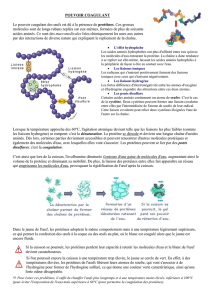
Le tableau périodique des éléments, ou tableau de Mendeleïev, comporte les 118 éléments
chimiques connus, ordonnés par numéro atomique croissant et organisés en fonction de leur
configuration électronique1. Chaque ligne du tableau est appelée "période" et correspond à
une couche électronique (c'est le nombre quantique principal, noté n). Il y a sept couches
électroniques connues à l'état fondamental, donc sept périodes dans le tableau périodique,
numérotées de 1 à 7. Chaque période comporte différentes orbitales atomiques, identifiées par
leur nombre quantique secondaire (noté l). Quatre types d'orbitales atomiques sont connues à
l'état fondamental (notées s, p, d et f et pouvant contenir chacune respectivement 2, 6, 10 et 14
électrons).
A partir du numéro atomique 104 on trouve les éléments chimiques synthétiques. Ce sont des
éléments instables avec une durée de vie si courte (de la microseconde au million d'années),
par rapport à l'âge de la Terre, que tous les atomes de ces éléments se sont désintégrés. Ils
peuvent cependant être recréés dans des accélérateurs de particules et dans les réacteurs
nucléaires.
Les colonnes du tableau périodique sont appelées "groupes". Il y a 18 groupes dans le tableau
périodique standard. A cela il faut ajouter les lanthanides et les actinides. Chaque groupe
constitue un ensemble d'éléments aux propriétés souvent bien distinctes des autres groupes.
Certains groupes ont des noms bien précis :
Groupe 1 : les métaux alcalins (excepté l'hydrogène)
Groupe 2 : les métaux alcalino-terreux
Groupe 17 : les halogènes
Groupe 18 : les gaz nobles (ou rares)
On utilise également d'autres noms pour regrouper les éléments dans le tableau de
Mendeleïev. Par exemple :
Métaux de transition : Il s'agit des éléments des périodes 4 à 7 et des groupes 3 à 12 (Sc, Ti,
V, Cr, Mn, Fe, etc.).
Métaux pauvres : Al, Ga, In, Sn, Tl, Pb, Bi. Ces métaux sont souvent mous (excepté Al), et
ont une température de fusion (et d'ébullition) inférieure à celles des métaux de transition.
Leur électronégativité est également plus élevée. Il n'existe pas de définition stricte d'un métal
pauvre.
1 Au-delà du numéro atomique 92, qui correspond à l'uranium, on entre dans le monde des
éléments artificiels instables (à courte durée de vie).

5
Métalloïdes : B, Si, Ge, As, Sb, Te, Po. Leurs propriétés physiques et chimiques sont
intermédiaires entre celles des métaux et des non-métaux. Il n'existe pas de définition stricte
d'un métalloïde.
Non-métaux : H, C, N, O, P, S, Se.
Les organismes vivants sont composés essentiellement de non-métaux (C, H, O, N, P, S).
Cependant, certains métaux alcalins (Na, K), certains alcalino-terreux (Ca, Mg) et certains
métaux de transition (Fe, Mn, Cu, Zn) sont indispensables au bon fonctionnement des
organismes. Pour les halogènes, Cl et F sont également retrouvés dans les organismes.
Les éléments chimiques sont identifiés dans le tableau périodique par leur numéro atomique,
qui représente le nombre de protons que contient leur noyau. Pour un même élément
chimique, et donc un même nombre de protons, le nombre de neutrons peut varier. Ces
atomes, qui occupent la même case dans le tableau périodique, sont appelés "isotopes".
Numéro atomique (Z) = Nombre de protons du noyau = Nombre d’électrons orbitant autour
du noyau (atome neutre).
Masse atomique (A) = Nombre de protons et de neutrons dans le noyau.
A = Z + N
N = Nombre de neutrons
Masse d’un atome ≈ A (électrons négligeables)
1.2.#Quantité#de#matière#
Pour calculer une quantité précise de matière on utilise la masse atomique (A).
Exemple : si on pèse 55,847 g de fer on obtiendra exactement 1 mole, c’est-à-dire 6,022 x
1023 atomes (nombre d’Avogadro).
Une mole de n'importe quel élément contient toujours le même nombre d’atomes.
Représentation habituelle : 12C et 56Fe
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
42
43
44
45
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
94
95
96
97
98
99
100
101
102
103
104
105
106
107
108
109
110
111
112
113
114
115
116
117
118
119
120
121
122
123
124
125
126
127
128
129
130
131
132
133
134
135
136
137
138
139
140
141
142
143
144
145
146
1
/
146
100%