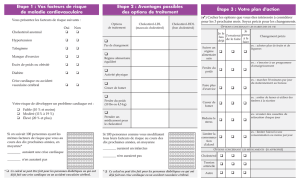
METABOLISME du CHOLESTEROL

. METABOLISME
DU CHOLESTEROL
A. GENERALITES
C’est un composé
polycyclique à 27 C
avec une fonction alcool « ol »
chole : par rapport au système biliaire
stérol : référence au noyau stérane
= perhydrocyclopentanophénentrène
Le phénentrène est composé de 3 noyaux
benzèniques accolés. Il s’y associe un cy-
clopentane, une chaîne latérale et des radi-
caux méthyles. La fonction alcool lui con-
fère une discrète hydrophilie, compensée par
les 27 C.
c’est un lipide, c'est à dire insoluble dans
l’eau
soluble dans les solvants organiques.
En représentation spatiale : importance pour les interactions métaboliques du cholestérol et de ses
dérivés de la position de l’hydroxyle et de la configuration des cycles A et B. La forme chaisedes
cycles A et B est la plus stable
A et B sont coplanaires.
Les produits dérivés du cholestérol (stéroïdes) seront à considérer du point de vue de leur encom-
brement.
B. ROLE DU CHOLESTEROL
1. Membranes cellulaires
a) constituant membranaire.
Le cholestérol est spécifique du monde animal. Il existe des stérols de structure voisine chez les
végétaux : phytostérols.
Le cholestérol est un constituant essentiel des membranes : jusqu’à 30 % des lipides membranaires
(érythroyctes notamment).
Il participe à l’isolement physique et électrique du compartiment intracellulaire.
1
HO
356
7
8
9
10
11
12 13
14 15
16
17
20
19
21 22
23
24 25 26
27
A B
C D
H3C
H3C
CH3
HO ABD
C

Il module la fluidité des membranes et est donc essentiel dans tous les transports membranaires.
La membrane cellulaire est une bicouche de phospholipides, de composition variable.
zone riche en AG saturés : risque de cristallisation, empêchée par le cholestérol (il fluidi-
fie).
zone riche en AG insaturés : tendance à l’excès de fluidité. A ce niveau, le cholestérol ri-
gidifie. b) transport plasmatique des lipides
Le cholestérol est un constituant essentiel du système de transport des lipides dans le plasma. Il
constitue en association avec des apoprotéines spécifiques des édifices macromoléculaires, les lipo-
préteines essentielles dans le transport des lipides dans le milieu aqueux que constitue le plasma.
permet le transport de quantités importantes de triglycérides (substances énegétiques pour les
organes). 2. Le cholestérol, précurseur de composés importants
a) acides biliaires
sécrétés en quantités importantes
émulsification des AG : agents tensioactifs permettant la digestion de graisses. la transformation
du cholestérol permet son élimination par les sels biliaires (1 g / jour environ).
b) vitamine D
La vit D (dihydrocholecaciférol) intervient dans le métabolisme phospho-calcique en régulant la
fixation du calcium sur la matrice osseuse. Elle est synthétisée à partir du cholestérol grâce aux UV.
(carence rachitisme).
c) hormones stéroïdes
Elles ont les cycles A B C D. La chaîne latérale est en général raccourcie (moins de 27 C).
Leur synthése se fait dans les gonades (hormones sexuelles) et les surrénales (cortisol...)
3. Les besoins en cholestérol
sont de environ 1 g par jour. Il viennent en partie de la synthèse endogène de novo et en partie de
l’alimentation (50 %, surtout dans les pays développés).
4. Cholestérol et pathologie
Il existe des relations entre nutrition, cholestérlémie et survenue de maladies cardio-vasculaires par
athérosclérose : dépots de composés lipidiques au niveau de l’intima des vaisseaux, ce qui diminue
leur calibre et rend insuffisante la quantité d’oxygène délivrée aux tissus (coeur...)
13 prix Nobel sont concernés par cette molécule.
C. BIOSYNTHESE
Sans membrane, pas de cellule ; sans cellule, pas de vie.
La synthèse est ubiquitaire, ce qui n’est pas surprenant quand on considère le caractère essentiel du
cholestérol.
Elle se produit majoritairement an niveau
du foie et de l’intestin
des organes ayant des fonctions spécifiques comme les gonades, les surrénales qui ont des fonctions
très actives par rapport à leur masse.
Elle se fait surtout à partir d’acétate activé : l’acétyl-CoA (2 C).
L’utilisation de précurseurs radioacifs marqués sur le méthyl :
ont produit le produit suivant :
H3C C
O
O-
*

Si c’est l’autre carbone (carbonyle) qui est marqué, on obtient la radioactivité complémentaire.
Actuellement, on dispose de produits pouvant réguler la biosynthèse du cholestérol.
K Bloch (1947) a montré la filiation entre acétate et cholestérol :
L’étape clef d’engagement dans la biosynthèse du cholestérol est la formation du composé en C6.
on agit sur cette étape pour réguler la biosynthèse du cholestérol.
C5 est l’élément monomèrique essentiel pour la biosynthèse ultérieure. Il est très voisin de
l’isoprène. 1. Formation du mévalonate
C’est l’étape d’engagement.
Condensation d’acétoacétyl-CoA avec de l’acétyl-Coa : sous l’action d’une enzyme très spécifique :
hydroxyméthylglutaryl synthétase (HMG synthase)
NB : le produit à l’origine du mévalonate (HMG-CoA) peut avoir 2 destinées différentes :
dans le cytosol ou le reticulum endoplasmique : voie biosynthétique cholestérol.
dans les mitochondries : on a une HMG lyase qui donne de l’acétyl-CoA et de l’acétoacétate qui
est le chef de file des corps cétoniques.
suivant le compartiment cellulaire on suit soit une voie anabolique soit une voie catabolique.
*
*
*
**
**
*
*
**
*
*
*
C2C6C5
C5
C10
C5C15
C15
C30 C27
acétate mévalonate
isopentényl PP
condensation
CO2
décarboxylation
condensation
condensation
géranyl PP
farnésyl PP
squalène
condensation

Dans la voie biosynthétique intervient secondairement une réductase qui en présence du cofacteur
NADPH + H+, va conduire au mévalonate.
Cette réductase est la cible de la régulation physiologique et pharmacologique (si maladie génétique
fréquente).
2. Transformation du mévalonate C6 C5
Elle nécessite une activation du mévalonate par 3 phosphorylations successives qui vont agir sur les
2 hydroxyles formation d’un groupement P-P et d’un groupement P sur le C3. Ce composé tri-
phosphorylé est quasiment virtuel et se décarboxyle facilement avec perte du CO2 en 1 pour former
l’isopentenyl-pyrophosphate.
H3CC~S-CoA
O
H3CC~S-CoA
OCH2C~S-CoA
O
CH3
O
C
HS-CoA
acétyl CoA
acétoacétyl CoA
HS-CoA
acétyl CoA
CH2
C~S-CoA
CH2
OH CCH3
COO-
O
3-hydroxy-3-méthylglutaryl CoA
HMG-CoA synthase
thiolase
CH2
CH2OH
CH2
OH CCH3
COO-
mévalonate
HS-CoA
2 NADPH + 2 H+
2 NADP+
HMG-CoA réductase
réaction clef
CH2
CH2OH
CH2
OH CCH3
COO-
mévalonate
3 ATP 3 ADP CH2
CH2O- P ~ P
CH2
P -O C CH3
COO-
ATP nº 1 ATP nº 2
ATP nº3
CH2
CH2O- P ~ P
CH2
CCH3
CO2 + Pi
isopentényl-
pyrophosphate
kinases
pyrophospho-
mévalonate
décarboxylase

L’isopentényl pyrophosphate est très proche de l’isoprène : 2-méthyl-1,3-butadiène. Très répandu
dans le monde animal et végétal. Il donne par polymérisation un dérivé synthétique proche du
caoutchouc
L’isopentényl-PP peut s’isomériser aisément, en présence d’une isomérase spécifique, en diméthy-
lallyl-PP :
3. Condensations
La condensation d’un diméthylallyl PP avec un isopentényl PP conduit, en présence d’une prényl
transférase, au géranyl PP :
La prényl transférase permet d’ajouter un nouvel isopentényl PP : farnésyl
H2CC
CH2
H3C
CH2O- P ~ P
isopentényl-
pyrophosphate
H2CC
CH
H3C
CH2
H3CC
CH
H3C
CH2O- P ~ P
diméthylallyl-
pyrophosphate
H2CC
CH2
H3C
CH2O- P ~ P
isopentényl-
pyrophosphate
géranyl-pyrophosphate
H3CC
CH
H3C
CH2O- P ~ P
diméthylallyl-
pyrophosphate
CH2O- P ~ P
prényl
transférase
P ~ P
 6
7
8
9
10
11
12
13
14
6
7
8
9
10
11
12
13
14
1
/
14
100%