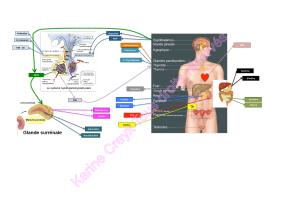
Mécanismes d`action de l `aldostérone dans le rein - iPubli

SYNTHÈSE
Directeur de Recherche Inserm
Prosseur l'Université de
Lausanne
Directeur de Recherche Inserm
J.-P.
L
I,
II,
I.
I
II

à
à

T
E

1
II

mfs
 6
7
6
7
1
/
7
100%