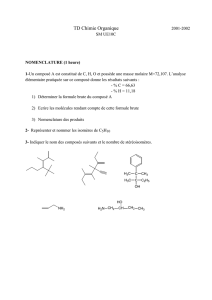
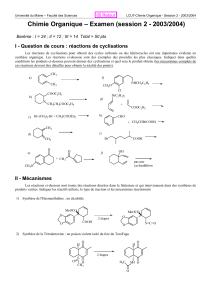
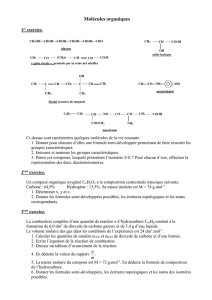
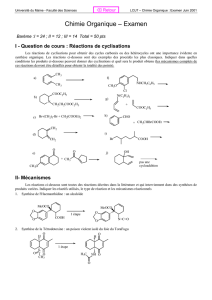
Coenzymes et Vitamines

Coenzymes et Vitamines
Introduction – définitions :
1). Coenzyme : cofacteur nécessaire pour qu’une enzyme devienne fonctionnelle.
L’enzyme est alors présente sous 2 états :
• Apoenzyme : enzyme SANS son coenzyme
• Holoenzyme : enzyme associée à son coenzyme
Il existe 2 types de coenzymes :
• Coenzyme co-substrat : subit une transformation lors de la réaction enzymatique :
S P
- 1 -
CoΣ CoΣ transformé
• Coenzyme co-facteur : présent dans le cœur catalytique de l’enzyme, non
transformé à la fin de la réaction.
2). Les vitamines :
déf. : Famille hétérogène de molécules organiques nécessaires au métabolisme, apportées par
l’alimentation, indispensables à la vie et sans valeur énergétique. Elles sont présentes à l’état de
traces dans l’alimentation.
Les aliments sont composés de nutriments :
• Les macronutriments avec valeur énergétique : Glucides, protides, lipides.
• Micronutriments : de nature organique (vitamines) et minérale (oligo-éléments).
C’est pour cela qu’on ne considère pas les acides gras essentiels comme des vitamines car ce sont
des macronutriments (apportant de l’énergie et tout).
Babarnab-PROD. Cours du Pr. Jean-Louis Guéant.

a). Classification physico-chimique des vitamines :
* Vitamines liposolubles : A, D, E, K
* Vitamines hydrosolubles : B6, B1, B9, B12, B5 / B2, B3, C
b). Classification fonctionnelle :
* Vitamines pseudo-hormonales : Vit. A et E
* Vitamines à mode d’action coenzymatique :
• soit réaction de transfert d’électron (oxydoréduction),
• soit réaction de transfert de groupements carbonés.
c). Apports nutritionnels conseillés :
ANC = apports nécessaires pour couvrir les besoins de 95% de la population.
I. Les vitamines liposolubles :
1). La vitamine A :
a). Structure :
C’est un alcool isoprénique : rétinol.
Composé d’un cycle β-ionone et d’une chaîne isoprénique.
* Schéma de la forme All-TRANS rétinol :
CH3 CH3 CH3 CH3
CH2OH fonction alcool
+ Site de phosphorylation
- 2 -
site d’isomérisation
CH3
1
12
11
Babarnab-PROD. Cours du Pr. Jean-Louis Guéant.

Comme cette molécule est sensible aux U.V, sous l’effet de la lumière il y a isomérisation au
niveau des carbones 11 et 12. On obtient la forme 11-CIS rétinol :
CH3 CH3 CH3
- 3 -
CH2OH
CH3
CH3
12
11
1
La forme habituelle de cette vitamine est le rétinol, les autres formes sont appelées vitamères.
Vitamères = isoformes d’une vitamine (et non d’un coenzyme !)
Rétinol : - CH2OH
Rétinal : - CHO
Acide rétinoïque : - COOH
Rétinyl Phosphate : - CH2O – P – O-
O
O-
L’acide rétinoïque est la forme active principale.
On passe du rétinol au rétinal, puis à l’acide rétinoïque par oxydations successives en
intracellulaire.
* Les apports :
L’alimentation animale apporte du rétinol sous forme estérifiée avec des acides gras. Selon la
longueur de la chaîne on a :
• rétinyl acétate
• rétinyl palmitate
Dans les végétaux, on trouve le β-carotène. Il possède 2 structures β-ionones :
Plan de symétrie
Point de clivage par une DIOXYGENASE
15’
15
Babarnab-PROD. Cours du Pr. Jean-Louis Guéant.

Le β-carotène peut subir un clivage par la 15-15’ β-carotène dioxygénase.
Il peut subir des oxydations successives par le métabolisme intestinal pour aboutir au rétinol.
Le β-carotène est un équivalent rétinol : 6 µg de β-carotène ⇔ 1 µg de rétinol.
b). Propriétés physiques :
Soluble dans les solvants organiques, dans l’éther, le formol, l’éthanol …
Très sensible à l’oxydation et à la lumière.
c). Sources :
* Sources animales (lipides) : abats, foie de poisson ou de veau, huiles de poisson, beurre, oeufs ...
* Sources végétales : carottes, myrtilles, tomates ...
d). Métabolisme :
Présent sous forme de rétinyl-esters dans l’alimentation.
Les rétinyls-esters sont transportés dans le tube digestif sous forme de micelles, puis une estérase
hydrolyse les liaisons esters :
Rétinyl ester rétinol
- 4 -
Estérase
Transport sanguin dans les Chylo
Passage barrière intestinale
Re-esterification et incorporation
dans les chylomicrons
FOIE
Dans le foie, le rétinol est stocké dans les cellules de Ito sous forme de rétinyl ester.
Le rétinol peut être libéré, il sera alors associé à une protéine de transport spécifique synthétisée
par le foie : la RBP (« rétinol binding prot »).
De plus, le complexe RBP-rétinol va se lier à la Transthyrétine pour éviter de se faire éliminer
par le rein. (⇒ Le complexe RBP-rétinol seul fait 21000 Da, et le glomérule filtre toutes les
substances dont le poids moléculaire est inférieur à 65000 Da).
Au final, c’est tout le complexe RBP-tranthyrétine-rétinol qui sera reconnu par un récepteur
d’endocytose au niveau des cellules périphériques.
Une fois dans la cellule, le rétinol subit 2 oxydations successives qui aboutissent à la formation
d’acide rétinoïque (=RA).
Dans le cytoplasme, ce dernier est associé à la CRABP (= « cytoplasm RA binding prot »).
(⇒ constitution du pool cellulaire en acide rétinoïque ?)
Babarnab-PROD. Cours du Pr. Jean-Louis Guéant.

Il existe 2 récepteurs pour l’acide rétinoïque (= facteurs de transcription = récepteurs nucléaires) :
• Le RAR = récepteur pour l’ All-TRANS acide rétinoïque.
• Le RXR = récepteur pour le 9-CIS acide rétinoïque.
Lorsque le ligand se lie à son récepteur spécifique, il y a translocation dans le noyau puis
dimérisation avec un autre complexe ligand-récepteur, ce qui permet d’externaliser les sites de
fixation à l’ADN qui sont sous la forme de structures en doigts de zinc. (Structures s’intercalant
dans les sillons de l’ADN) :
Zn
2
+
* RAR : plusieurs types : RARα, RARβ, RARγ.
Ils s’associent ensemble pour former des hétérodimères.
* RXR : également 3 formes, mais il forme des hétérodimères avec d’autres récepteurs :
• Récepteur de l’hormone thyroïdienne
• Récepteur nucléaire de la vitamine D
• Récepteurs PPAR
L’effet final de la fixation sur l’ADN est d’assurer un rôle de transactivation : ces complexes
augmentent la transcription des gènes cibles. Ces gènes cibles sont nombreux et sont impliqués
dans le cycle cellulaire et la différenciation cellulaire.
note : application dans le traitement de mélanomes + cancers bronchiques ( ... ?).
* Structure générale d’un récepteur nucléaire :
pour translocation nucléaire
- 5 -
liaison du ligand
A B C E
D
Domaines transactivateurs domaine de liaison à l’ADN
Babarnab-PROD. Cours du Pr. Jean-Louis Guéant.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
24
25
26
27
28
29
30
31
32
33
34
35
36
37
38
39
40
41
1
/
41
100%