10/10/2014 PERREARD Pauline L3 SNP Dr Trebuchon-Da

!"#$!" % &'()* %
!!(+"#$ "%!' +,+(*%(+
''+&''()*+'' ($!)-
$+(.'(+/0('+1++ +/+)((+,"#$ -
/(12(+3/4(.())(-
"+'%
–#$ %!56(1)(-
–#$ %!!#,%!561)(-
+(() .5(+((
.'(6)(+%+)(-
(+1++3,(.+3/
)1)---/))(+()&(((1
'(()1') 3/+
()1)-7$/(/--,.//(/((
+,6)(3,8(+1+,6)(
('-,0)'+.('
+)((() -
"'++&( 11()-
"('+&( 9(()(+6)((&&(-
Plan
A. L'oeil
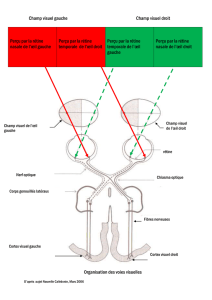
B. Transfert à travers les voies optiques centrales (voie rétino-géniculo-corticale)
C. Traitement de l'information corticale

/6)((+'.6).((&(+0(()((.)
2(+)')((+(.(+3(+1+661))(-:'
-!!-;
#63/6)('((6)1((+/('+$"/
'+ !('03/#$0#"$"1$<(((1./(3')((
+/'##-
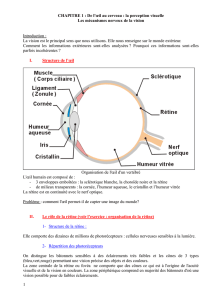
A. L'oeil : traitement de l'information au niveau rétinien.
/)(+(6))*:+1() 1(3;) '&:((+/(.;
'+1(./(,$!#-
'(+/'1(.)*1.((6-"
'+3((1/)+(1(-
!(+66+'333('+)113/(3
&(((2(*)+/((+(<(3/) (61)-

2)#$"!#$'+16(+&3,$76(+
/6)()-! "#$
%"#$
3)4$')((,#!&+661("! 5$-!((1(
1(((())1++&(2+6%
–))7+((+7)2+ 9&6+
–+)(17((+7)2 9&6
((1/6)(+.(+)5/1-
)+/((=15 $!0""-%2(*)$ ,%!
)2+3,$ ,%!)()2-
6)#$(1 )(#$.(5+661+1/(5
%!-!22(*)+)+6(++(.,## -!0(23
,$(=)0+,$5-
7+2(2(+((1+(.2++/))+(&(
'+(('+*-
*+ ( $6#!#%
–(+(%+,$"#,!$:=(9((;
–()2%#!1,$"cellules horizontales&$
–(&):>+)*;%#!$(+1(+&'6
(3-
$/( )(
&&+
3+(
6(3-
?(&(+
1(('
71(1) )(3
0=+
((+(
(+(1(-

6'1((1(+)(:(0;-
+((+1(+1(,()<)((-
–!/2+9(('+6'1.1(((=(+1(113-
–6'1.2(()&)+=.,(87#!" 0 79:
;#=)(((+'#(9(((5)(1-
$((((+=)('(*11 +-
& '
()'113(+1()1%
7@(%=7>>
7@113%)')(()(1)-
(')##&*#&*
!23,,$"#,!$%=(9((-
$(2++66*(+(()+
(()(((15*+)<)64:&
,$;%
– )(&()(( , %!+
3(1,,
– )(('#$,#!$!$
,%!:2';
1(((+(1(%

<1=2'''-,!1*! 5$%!#>:!(+'
#,%!:+(;-!2)A-B+(+()*11(
+66++661-+,
=(+',,%!:0;-!)(((+ +((
163+( +/++-!2C)A-
,, (1*!!!$7,$#!5$:
!2( +(2+='+ )(+661(.11+'+:.'(
( ;2D(+=+(163)(/ -
-...
$!#E(+(+/1 ):1() 1(3;'(+((+
))'+(1(-(((%/2+5((1(5F((+/(
1115/1('(-
(&/ %
(6)(($=#!,, .3(/(++&)1%(
1(:1(*;-,?$"5@$,:
1(*&(+&6)%6)('(6)('-
$/((()+6((11)33')((++/1'*)('5
((+(-+)((1(+((/6)(-
;()&'
+-1(*
+6)56)(-!2
+)+6(+(+
++3('
)<)(1G%
$!#:
;(+()
,,"$(A):
;2+2GH
GH-((GH
++)-
;6+(1(.2+#!0%!B#<",-3(&+B#
!.&>+ )(&(6)(7(8(,$,$-
! 5$,$ -$ !$#!0%!4,$,$#!CC
A l'obscurité
)(&(+(1(.&>('((2(1((+> 9
56(((+GH-$(",$,$)(((5+ +
((+))(&(+<&' D- )((26(+()
'/&(-
EI6&((+>((' )(&(()16&+J>((+ )(
(-$+&)')(31(((1#!$71#!$":
K
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
1
/
20
100%