Cinétique Michaelienne - Pages Persos Chez.com

C
C
Ci
iin
n
né
é
ét
tti
iiq
q
qu
u
ue
e
eM
M
Mi
iic
c
ch
h
ha
a
ae
e
el
lli
iie
e
en
n
nn
n
ne
e
e
L'étude de la cinétique enzymatique nous permet d'identifier et de décrire les
mécanises des réactions biologiques et les mutations des enzymes en étudiant leur
vitesse réactionnelle.
On peut comprendre alors comment les modifications de l'environnement de
l'enzyme peuvent affecter son fonctionnement.
I
I_
_D
Dé
ét
te
er
rm
mi
in
na
at
ti
io
on
nd
de
el
l'
'a
ac
ct
ti
iv
vi
it
té
ée
en
nz
zy
ym
ma
at
ti
iq
qu
ue
e
A
A.
.V
Vi
it
te
es
ss
se
ei
in
ni
it
ti
ia
al
le
e
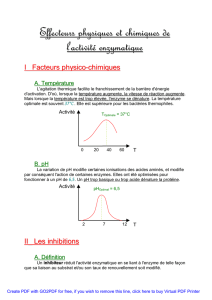
C'est la vitesse au début de la réaction de transformation du substrat en
produit par les enzymes, c'est-à-dire lorsque seulement moins de 10% de substrat
est consommé ( [S] Ycte ). Elle est caractéristique de l'enzyme.
B
B.
.C
Co
on
nc
ce
en
nt
tr
ra
at
ti
io
on
ne
en
ns
su
ub
bs
st
tr
ra
at
t
C
C.
.D
Dé
ér
ro
ou
ul
le
em
me
en
nt
td
de
el
la
ar
ré
éa
ac
ct
ti
io
on
n
Celle-ci s'effectue en 3 étapes :
[P]
t
État
préstationnaire
Plateau
État
stationnaire
Lors de l'état stationnaire, la concentration
en produit évolue linéairement et la pente
de la droite donne la vitesse initiale.
t
[P]
On quitte les conditions
de Michaelis :
[S] >> [E]0
[S]1
[S]2
[S]3vi
[S]
[S]1
[S]2
[S]3
vmax
vmax/2
KM
Create PDF with GO2PDF for free, if you wish to remove this line, click here to buy Virtual PDF Printer

I
II
I_
_I
In
nt
te
er
rp
pr
ré
ét
ta
at
ti
io
on
nd
de
eM
Mi
ic
ch
ha
ae
el
li
is
s
A
A.
.C
Ci
in
né
ét
ti
iq
qu
ue
e
l
lÉ
Éq
qu
ui
il
li
ib
br
re
er
ra
ap
pi
id
de
e
La cinétique nous donne :
or
On a donc la constante de dissociation :kD= [E][S] / [ES] = k-1 / k1
l
lÉ
Ét
ta
at
ts
st
ta
at
ti
io
on
nn
na
ai
ir
re
e
Dans les conditions de Michaelis ([S] >>[E]), l'approximation de l'état quasi-
stationnaire pour le complexe ES nous donne :
B
B.
.É
Éq
qu
ua
at
ti
io
on
nd
de
eM
Mi
ic
ch
ha
ae
el
li
is
s
On obtient la constante de Michaelis :kM= [E][S] / [ES] = (k-1+k2) / k1
Soit [ET] = [E] + [ES], alors on a
Remarque : lorsque la solution est saturée en substrat, [ET] = [ES] et vi= vmax.
l
lC
Co
ou
ur
rb
be
ed
de
eM
Mi
ic
ch
ha
ae
el
li
is
s
àAinsi pour [S] = kMon a vi= vmax/2
C
C.
.O
Ob
bt
te
en
nt
ti
io
on
nd
de
es
sp
pa
ar
ra
am
mè
èt
tr
re
es
s
q
qAjustement des données à l'équation de Michaelis (par ordinateur)
q
qObtention de vmax et kM
E + S k1
k-1ES
E + S k1
k-1ES k2E + P
vi
[S]
vmax
vmax/2
kM
Create PDF with GO2PDF for free, if you wish to remove this line, click here to buy Virtual PDF Printer

q
qLinéarisation par une double réciproque : 1/vi= f(1/[S])
q
qOn obtient 1/vmax et -1/kM
D
D.
.R
Re
ep
pr
ré
és
se
en
nt
ta
at
ti
io
on
nr
ré
ée
el
ll
le
ed
de
es
sp
pa
ar
ra
am
mè
èt
tr
re
es
s
On appelle k2le turn over, qui représente le nombre de cycles catalytiques
par seconde. Plus k2est élevée, plus l'enzyme est efficace.
On appelle kMl'affinité, qui représente la facilité qu'a un substrat de se lier à
une enzyme. Plus kMest petit, plus le substrat se liera facilement à l'enzyme.
On définit alors k2/kM( ou vmax/kM), qui permet de déterminer si la réaction
sera favorable et efficace entre l'enzyme et le substrat. Plus cette valeur est élevée,
plus la réaction est favorable.
I
II
II
I_
_R
Ré
éa
ac
ct
ti
io
on
ns
sa
av
ve
ec
ce
ef
ff
fe
ec
ct
te
eu
ur
rl
li
in
né
éa
ai
ir
re
e
On considère F, un effecteur linéaire. Il peut se lier à l'enzyme sur un site
unique, en activant ou en inhibant l'activité enzymatique.
Le mécanisme réactionnel sera de ce type :
àOn considère que la fixation de F sur l'enzyme est très rapide et que lors de
la mesure, les fixations ont atteint l'équilibre.
èOn utilisera les constantes d'équilibre et non les constantes de vitesse de toutes
les réactions impliquant F.
A
A.
.I
In
nh
hi
ib
bi
it
ti
io
on
n
l
lC
Ci
in
né
ét
ti
iq
qu
ue
e:
:
E + S
+ F
k1
k-1ES
+ F
k2E + P
EF
+ S EFS E + P + F
ak2
Si activateur
KFK'F
E + S k1
k-1ES k2E + P kM= [E][S] / [ES]
- 1/KM
1/vmax
1/vi
1/[S]
Pente
KM/vmax
Create PDF with GO2PDF for free, if you wish to remove this line, click here to buy Virtual PDF Printer

l
lC
Co
om
mp
pé
ét
ti
it
ti
io
on
n:
:
On distingue alors plusieurs cas :
q
qInhibition compétitive : la fixation de l'inhibiteur sur l'enzyme empêche la
fixation du substrat et inversement ( KI<< K'I)
k'M= kMx ( 1 + [I] / kI)
q
qInhibition non compétitive : la fixation de l'inhibiteur sur l'enzyme
n'empêche pas la fixation du substrat, mais empêche sa transformation ( KI
= K'I)v'max = vmax / ( 1 + [I] / kI)
q
qInhibition incompétitive : l'inhibiteur ne se lie qu'au complexe
enzyme/substrat ( K'I<< KI)
k'M= kM/ ( 1 + [I] / k'I)
v'max = vmax / ( 1 + [I] / k'I)
q
qInhibition mixte : ( K'I@KI)
k'M= kMx [( 1 + [I] / kI) / ( 1 + [I] / k'I)]
v'max = vmax / ( 1 + [I] / k'I)
l
lN
Na
at
tu
ur
re
ed
de
es
si
in
nh
hi
ib
bi
it
te
eu
ur
rs
s:
:
Dans le cas des inhibiteurs compétitifs, ceux-ci peuvent :
àPrésenter une similarité structurale avec la substrat
àÉtablir le même type d'interactions avec l'enzyme
àPrésenter une analogie avec un état de transition
Ex : l'AZT pour traiter le SIDA : c'est un inhibiteur de la transcriptase inverse
àIl se substitue à la thymine
B
B.
.A
Ac
ct
ti
iv
va
at
te
eu
ur
rs
s
On observe plusieurs cas, selon que l'activateur et le substrat se fixent au
hasard ou d'une manière séquentielle à l'enzyme.
1 / vi
1/ [S]
-1/kM-1/k'M
1 / v'max
Sans inhibition
Inhibition non
compétitive
Inhibition
compétitive
Inhibition
incompétitive
E + I EI KI= [E][I] / [EI]
ES + I EIS K'I= [ES][I] / [EIS]
Create PDF with GO2PDF for free, if you wish to remove this line, click here to buy Virtual PDF Printer

q
qFixation indépendante : la fixation des deux substrats se fait sur des sites
différents, et la fixation de l'un ne modifie pas la fixation de l'autre ( KA= K'A
et KS= K'S)
v'max = vmax / ( 1 + KA/ [A] )
k'M= KS
q
qFixation dépendante : la fixation d'un des ligands affecte l'affinité de
l'enzyme pour l'autre ligand ( KA@K'Aet KS@K'S)
k'M= K'Sx [( 1 + KA/ [A]) / ( 1 + K'A/ [A])]
v'max = vmax / ( 1 + K'A/ [A] )
q
qFixation séquentielle :
q
qL'activateur se lie en premier, puis vient le substrat.
k'M= KSx ( 1 + KA/ [A] )
q
qLe substrat se lie en premier, puis vient l'activateur.
àOn obtient :
I
IV
V_
_E
En
nz
zy
ym
me
es
sà
àd
de
eu
ux
xs
su
ub
bs
st
tr
ra
at
ts
s
E + S
+ A
KSES
+ A
EA
+ S EAS E + P + A
kcat
KAK'A
K'S
KS= [E][S]/[ES] K'S= [EA][S]/[EAS]
KA= [E][A]/[EA] K'A= [ES][A]/[EAS]
Et KSK'A= K'SKA
E + S
+ A
KAEA + S kcat
EAS
KSEA + P
E + S
+ A
KA
ES + A kcat
EAS
KSEA + P
1 / vi
1/ [S]
-1/kM
1 / vmax Activation
indépendante
Sans activation
Activation
séquentielle
Create PDF with GO2PDF for free, if you wish to remove this line, click here to buy Virtual PDF Printer
 6
7
8
9
6
7
8
9
1
/
9
100%