PSY 1055. Psychologie de la perception. Bases physiologiques de

1
PSY 1055. Psychologie de la
perception.
Bases physiologiques de la
perception visuelle, II.
Frédéric Gosselin /
Eric McCabe
Plan du cours
• On résume et termine (au besoin) le cours de la
semaine dernière…
• On poursuit l’étude de la physiologie du
système visuel
– Corps genouillé latéral (CGL/LGN).
– Cortex visuel primaire
lumière
Corps genouillé
latéral (LGN) gauche
Corps genouillé
latéral (LGN) droit
Cortex visuel
Stimulus et lumière
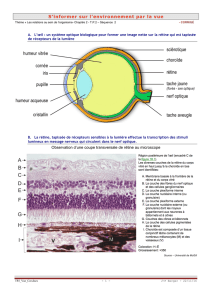
Œil et rétine
Retour sur la semaine dernière
1)Focalisation,
2)Degré d’angle visuel,
tan(α/2) = (l/2)/d
3)Rétine : Types de
cellules,
4)Rétine : Transduction,
5)Adaptation à la noirceur
6)Rétine : Convergence
1)Amplitude*sin(fréquence*angle+phase)
2)Luminance
1 s0 s
sin(2*angle) sin(angle+.39)
2*sin(angle)
sin(angle)
.39 s

2
lumière
Corps genouillé
latéral (LGN) gauche
Corps genouillé
latéral (LGN) droit
Œil et rétine
Retour sur la semaine dernière
7)CR des cellules ganglionnaires (Vaughan)
- Concentriques
- Antagonisme centre-périphérie
8)Rétine : Inhibition latérale
9)Illusions (Hermann, Contraste simultanée,
Mach, White)
10)Scotome
Pathologies : 1)Focalisation, 2)Médium
oculaire, 3)Rétine, 4)Nerf optique
lumière
Corps genouillé
latéral (LGN) gauche
Corps genouillé
latéral (LGN) droit
2- Cortex visuel
Plan du cours
1- Corps
genouillés
latéraux
Traitement de l’information dans le CGL
Jouerait un rôle dans la régulation de l’information en provenance de la rétine.
+
10 nerfs de la rétine →
4 nerfs vers le cortex
Vers cortex
En plus de recevoir les signaux optiques, le CGL reçoit des entrées en provenance
du cortex visuel, du tronc cérébral, d’autres noyaux thalamiques et d’autres
neurones du CGL.
Du cortex
De la rétine
LGN
Thalamus-LGN
Tronc cérébral
=

3
Les champs récepteurs du CGL
Le champs récepteur d’un neurone du CGL est une région sur la
rétine qui, lorsque stimulée, permet le déclenchement de ce
même neurone du CGL (Goldstein, 2001)
= luminance haute
= luminance basse
Le fond de la rétine
+
Champs récepteur d’une cellule du LGN
(“center-surround”)
• Les champs récepteurs des neurones du CGL ont la même
configuration concentrique que les cellules ganglionnaires.
Il existe aussi des champs récepteurs “center OFF-surround ON” dans le LGN.
(Ozhawa, DeAngelis & Freeman 1995)
Organisation dans le LGN
(couches)
Chaque LGN a 6 couches (la couche 1 étant la plus
profonde et 6, la plus superficielle).
Oeil ipsilatéral : couches 2, 3 et 5
Oeil controlatéral : couches 1, 4 et 6

4
Organisation dans le LGN
(rétinotopie)
Chaque couche d’un LGN est rétinotopique. Les neurones
situés près les uns des autres dans une structure
cérébrale ont des champs récepteurs correspondant à des
localisations rétiniennes voisines.
Oeil
Organisation dans le LGN
(magno vs parvo)
• Les couches 1 et 2 sont magnocellulaires. C’est-à-
dire qu’elles reçoivent des axones de cellules
ganglionnaires de type M).
• Les autres couches sont parvocellulaires. C’est-à-
dire qu’elles reçoivent des axones de cellules
ganglionnaires de type P).
Organisation dans le LGN
(magno vs parvo II)
• Les cellules des couches magnocellulaires
et parvocellulaires ont des rôles différents
dans la perception visuelle.
• Lésions des couches magno :
– Atteinte de la perception du mouvement.
• Lésions des couches parvo :
– Atteinte de la perception des couleurs, de la texture, de
la profondeur et des contours.

5
Plan pour le reste du cours
lumière
Corps genouillé
latéral (LGN) gauche
Corps genouillé
latéral (LGN) droit
Cortex visuel
primaire (V1)
Régions de traitement du signal
visuel dans le cortex
Cortex visuel
primaire (V1) ou
cortex strié
Cortex extrastrié
Cortex visuel primaire
• Au contraire des neurones du CGL, ceux du cortex
strié ne répondent pas particulièrement bien à des
points lumineux.
 6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
1
/
23
100%