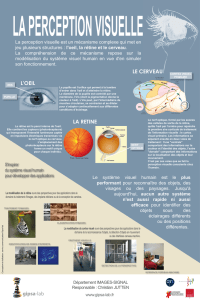
PSY 1055. Psychologie de la perception. Bases physiologiques de

1
PSY 1055. Psychologie de la
perception.
Bases physiologiques de la
perception visuelle, II.
Frédéric Gosselin /
Eric McCabe
lumière
Corps genouillé
latéral (LGN) gauche
Corps genouillé
latéral (LGN) droit
Cortex visuel
Stimulus et lumière
Œil et rétine
Retour sur la semaine dernière
1)Focalisation,
2)Degré d’angle visuel,
tan(α/2) = (l/2)/d
3)Rétine : Types de cellules,
4)Rétine : Transduction,
5)Adaptation à la noirceur
6)Rétine : Convergence
1)Amplitude*sin(fréquence*angle+phase)
2)Luminance
0 1 2 3 4 5 6 7
-2
-1.5
-1
-0.5
0
0.5
1
1.5
2
1 s0 s
sin(2*angle) sin(angle+.39)
2*sin(angle)
sin(angle)
.39 s
lumière
Corps genouillé
latéral (LGN) gauche
Corps genouillé
latéral (LGN) droit
Œil et rétine
Retour sur la semaine dernière
7)CR des cellules ganglionnaires (Vaughan)
- Concentriques
- Antagonisme centre-périphérie
8)Rétine : Inhibition latérale
9)Illusions (Hermann, Contraste simultanée,
Mach, White)
10)Scotome
Pathologies : 1)Focalisation, 2)Médium
oculaire, 3)Rétine, 4)Nerf optique
Scotome : tache aveugle pathologique
• Limité à un oeil : lésion au
niveau de la rétine ou du
nerf optique
• Dans les deux hémichamps
temporaux (mais pas
nasaux) : lésion au niveau
du chiasme optique
• Dans les deux hémichamps
gauches (ou droits) : lésion
au niveau de tractus optique
lumière
Corps genouillé
latéral (LGN) gauche
Corps genouillé
latéral (LGN) droit
2- Cortex visuel
Plan du cours
1- Corps
genouillés
latéraux
Attention : erreur
dans le codex
Traitement de l’information dans le CGL
Jouerait un rôle dans la régulation de l’information en provenance de la rétine.
+
10 nerfs de la rétine →
4 nerfs vers le cortex
Vers cortex
De la rétine
LGN
En plus de recevoir les signaux optiques, le CGL reçoit des entrées en provenance
du cortex visuel, du tronc cérébral, d’autres noyaux thalamiques et d’autres
neurones du CGL.
Du cortex
Thalamus-LGN
Tronc cérébral
=

2
Les champs récepteurs du CGL
Le champs récepteur d’un neurone du CGL est une région sur la
rétine qui, lorsque stimulée, permet le déclenchement de ce
même neurone du CGL (Goldstein, 2001)
= luminance haute
= luminance basse
Le fond de la rétine
+
Champs récepteur d’une cellule du LGN
(“center-surround”)
• Les champs récepteurs des neurones du CGL ont la même
configuration concentrique que les cellules ganglionnaires.
Il existe aussi des champs récepteurs “center OFF-surround ON” dans le LGN.
(Ozhawa, DeAngelis & Freeman 1995)
Organisation dans le LGN
(couches et oeil d’origine)
Chaque LGN a 6 couches (la couche 1 étant la plus
profonde et la 6 la plus superficielle).
Oeil ipsilatéral : couches 2, 3 et 5
Oeil controlatéral : couches 1, 4 et 6
Organisation dans le LGN
(rétinotopie)
Chaque couche d’un LGN est rétinotopique. Les neurones
situés près les uns des autres dans une structure cérébrale
ont des champs récepteurs correspondant à des
localisations rétiniennes voisines.
Oeil
Organisation dans le LGN
(magno vs parvo)
• Les couches 1 et 2 sont magnocellulaires. C’est-à-
dire qu’elles reçoivent des axones de cellules
ganglionnaires de type M).
• Les autres couches (3,4,5,6) sont parvocellulaires.
C’est-à-dire qu’elles reçoivent des axones de cellules
ganglionnaires de type P).
Attention : Erreur
dans le Goldstein,
table 3.1 page 94
Organisation dans le LGN
(magno vs parvo II)
• Les cellules des couches magnocellulaires
et parvocellulaires ont des rôles différents
dans la perception visuelle.
• Lésions des couches magno :
– Atteinte de la perception du mouvement.
• Lésions des couches parvo :
– Atteinte de la perception des couleurs, de la texture, de
la profondeur et des contours.

3
Plan pour le reste du cours
lumière
Corps genouillé
latéral (LGN) gauche
Corps genouillé
latéral (LGN) droit
Cortex visuel
primaire (V1)
Régions de traitement du signal
visuel dans le cortex
Cortex visuel primaire
(V1) ou cortex strié
Cortex extrastrié
Cortex visuel primaire
• Au contraire des neurones du CGL, ceux du cortex
strié ne répondent pas particulièrement bien à des
points lumineux.
Nobel en physiologie en 1981
D. Hubel T. Wiesel
Ozhawa, DeAngelis & Freeman
1995
Classique Corrélation inversée
Visionner fichiers Quicktime…
Origine des cellules simples?
Champs récepteurs
des cellules du LGN Champ récepteur
d’une cellule simple

4
Le champs récepteur
• Le champs récepteur d’un neurone du cortex strié est la région
sur la rétine qui, lorsque stimulée, a un effet [maximal] sur le
déclenchement de ce même neurone.
Le fond de la rétine
!!!
= peu importe
= luminance haute
= luminance basse
Organisation dans V1
Rétinotopie dans V1 révélée par
autoradiographie
• On montre cette image à un singe +
injection de sucre radioactif.
Rétinotopie dans V1 : Résultats
Magnification corticale : Variation de la surface corticale dédiée à la
représentation d’un stimulus en fonction de la localisation des récepteurs
qu’il stimule.
Stimulus Cortex
Explication
• Une plus grande surface corticale est donc allouée
à la représentation de la fovéa par rapport à la
périphérie.
• 0.01% des cellules de la rétine (les cellules de la
fovéa) atteignent 8-10% du cortex visuel.
• Facteurs de magnification corticale:
– (a) plus de cellules dans la fovéa par unité de surface et
–(b) plus grand nombre de cellules corticales dédiées à
chaque cellule de la fovéa.
Organisation dans V1
Pénétrations perpendiculaires
Pénétration oblique

5
Colonne d’orientation
• Les neurones rencontrés lors d’une pénétration perpendiculaire
(tracés A et B ci-bas) ont des champs récepteurs ayant la même
préférence à l’orientation (colonne d’orientation).
• Lors d’une pénétration oblique, l’orientation préférée varie de
façon continue.
Les hypercolonnes
• Module pour le traitement de la stimulation d’une
région rétinienne.
- Une hypercolonne comprend
deux colonnes de dominance
oculaire (oeil droit et gauche).
- Une hypercolonne
mesure environ 1 mm3.
perpendiculaire
oblique 1 hypercolonne =
1 colonnne de localisation =
2 colonnes de dominance oculaire =
2 ensembles des colonnes d’orientation
Rétinotopie
• L’ensemble des hypercolonnes est rétinotopique (c’est-à-
dire que 2 hypercolonnes adjacentes ont des champs
récepteurs adjacents sur les rétines). Mais il y a
grossissement à la fovéa.
• De plus, les champs récepteurs des cellules d’une même
hypercolonne occupent des positions correspondantes sur
les rétines.
Les hypercolonnes (suite)
• Chaque colonne de dominance oculaire possède 6
couches.
Le rôle des cellules simples
Orientation
Fréquence
Phase
Contraste
Rappelez-vous :
onde sinusoidale = Amplitude * sin(fréquence * angle + phase)
Ici on ajoute l’orientation
Relation avec les cellules simples
• En fait, les cellules simples sont sensibles à
trois attributs des stimulations rétiniennes
– La fréquence spatiale.
– l’orientation.
– et la position sur la rétine.
Démonstration
 6
7
8
9
10
11
6
7
8
9
10
11
1
/
11
100%